
Ботаника -Botany

| Часть серии о |
| Биология |
|---|
|
|
Ботаника , также называемая наукой о растениях , биологией растений или фитологией , является наукой о жизни растений и отраслью биологии . Ботаник , ботаник или фитолог — это ученый , который специализируется в этой области. Термин «ботаника» происходит от древнегреческого слова βοτάνη ( botanē ), означающего « пастбище », « травы », « трава » или « корм »; βοτάνη , в свою очередь, происходит от βόσκειν ( boskein ), «кормить» или « пастись ». Традиционно ботаника включала также изучение грибов и водорослей микологами и психологами соответственно , при этом изучение этих трех групп организмов оставалось в сфере интересов Международного ботанического конгресса . В настоящее время ботаники (в строгом смысле) изучают около 410 000 видов наземных растений , из которых около 391 000 видов — сосудистые растения (в том числе около 369 000 видов цветковых ), а около 20 000 — мохообразные .
Ботаника возникла в доисторические времена как травничество благодаря усилиям древних людей идентифицировать, а затем культивировать съедобные, лекарственные и ядовитые растения, что сделало ее одной из древнейших отраслей науки. Средневековые физические сады , часто при монастырях , содержали растения, имеющие медицинское значение. Они были предшественниками первых ботанических садов при университетах , основанных с 1540-х годов. Одним из первых был ботанический сад Падуи . Эти сады способствовали академическому изучению растений.
В 19 и 20 веках были разработаны новые методы изучения растений, в том числе методы оптической микроскопии и визуализации живых клеток , электронной микроскопии , анализа числа хромосом , химии растений , структуры и функции ферментов и других белков . В последние два десятилетия 20-го века ботаники использовали методы молекулярно-генетического анализа , включая геномику , протеомику и последовательности ДНК, для более точной классификации растений.
Современная ботаника — это обширный междисциплинарный предмет, в который внесены вклады и идеи из большинства других областей науки и техники. Темы исследований включают изучение структуры растений , роста и дифференциации, воспроизводства , биохимии и первичного метаболизма , химических продуктов , развития , болезней , эволюционных взаимоотношений , систематики и таксономии растений . Доминирующими темами в науке о растениях 21 века являются молекулярная генетика и эпигенетика , которые изучают механизмы и контроль экспрессии генов во время дифференцировки растительных клеток и тканей . Ботанические исследования имеют разнообразные применения в обеспечении основных продуктов питания , таких материалов, как древесина , масло , каучук, волокна и лекарства, в современном садоводстве , сельском и лесном хозяйстве , размножении растений , селекции и генетической модификации , в синтезе химикатов и сырья для строительства и производство энергии, управление окружающей средой и сохранение биоразнообразия .
История
Ранняя ботаника

Ботаника возникла как травничество , изучение и использование растений из-за их лечебных свойств. Ранняя зарегистрированная история ботаники включает множество древних писаний и классификаций растений. Примеры ранних ботанических работ были найдены в древних текстах из Индии, датируемых до 1100 года до н.
Современная ботаника уходит своими корнями в Древнюю Грецию , в частности, к Теофрасту (ок. 371–287 до н. Э.), Ученику Аристотеля , который изобрел и описал многие из ее принципов и широко известен в научном сообществе как «отец ботаники». Его основные работы « Исследование растений» и «О причинах возникновения растений » представляют собой наиболее важный вклад в ботаническую науку до средних веков , почти семнадцать столетий спустя.
Еще одна работа из Древней Греции, оказавшая раннее влияние на ботанику, — это De Materia Medica , пятитомная энциклопедия о фитотерапии , написанная в середине первого века греческим врачом и фармакологом Педанием Диоскоридесом . De Materia Medica широко читалась более 1500 лет. Важный вклад средневекового мусульманского мира включает « Набатейское земледелие » Ибн Вахшийи , « Книгу растений » Абу Ханифы Динавари (828–896) и « Классификацию почв » Ибн Бассаля . В начале 13 века Абу аль-Аббас ан-Набати и Ибн аль-Байтар (ум. 1248) систематически и научно писали о ботанике.
В середине 16 в. при ряде итальянских университетов были основаны ботанические сады . Ботанический сад Падуи в 1545 году обычно считается первым, который до сих пор находится на своем первоначальном месте. Эти сады продолжали практическую ценность более ранних «физических садов», часто связанных с монастырями, в которых выращивали растения для медицинских целей. Они поддерживали рост ботаники как академического предмета. Были прочитаны лекции о растениях, выращиваемых в садах, и продемонстрировано их медицинское применение. Ботанические сады появились в Северной Европе намного позже; первым в Англии был Ботанический сад Оксфордского университета в 1621 году. На протяжении всего этого периода ботаника оставалась прочно подчиненной медицине.
Немецкий врач Леонхарт Фукс (1501–1566) был одним из «трех немецких отцов ботаники» наряду с теологом Отто Брунфельсом (1489–1534) и врачом Иеронимом Боком (1498–1554) (также называемым Иеронимом Трагусом). Фукс и Брунфельс отошли от традиции копирования более ранних работ, чтобы сделать собственные оригинальные наблюдения. Бок создал свою систему классификации растений.
Врач Валериус Кордус (1515–1544) является автором важных с ботанической и фармакологической точки зрения книг о травах Historia Plantarum в 1544 году и фармакопеи непреходящего значения Dispensatorium в 1546 году. Натуралист Конрад фон Геснер (1516–1565) и травник Джон Джерард (1545–ок. 1611 ). ) опубликованы травники, посвященные лекарственному использованию растений. Натуралист Улисс Альдрованди (1522–1605) считался отцом естествознания , включавшего изучение растений. В 1665 году, используя первый микроскоп, эрудит Роберт Гук обнаружил клетки , термин, который он придумал, в пробке , а вскоре и в живой растительной ткани.
Ранняя современная ботаника

В течение 18-го века были разработаны системы идентификации растений , сравнимые с дихотомическими ключами , где неопознанные растения помещаются в таксономические группы (например, семейство, род и вид) путем выбора между парами признаков . Выбор и последовательность признаков могут быть искусственными в ключах, предназначенных исключительно для идентификации ( диагностические ключи ), или более тесно связаны с естественным или филетическим порядком таксонов в синоптических ключах. К 18 веку новые растения для изучения прибывали в Европу во все большем количестве из недавно открытых стран и европейских колоний по всему миру. В 1753 году Карл фон Линней (Carl Linnaeus) опубликовал свой Species Plantarum , иерархическую классификацию видов растений, которая остается точкой отсчета для современной ботанической номенклатуры . Это установило стандартизированную биномиальную или двухкомпонентную схему именования, в которой первое имя представляло род , а второе идентифицировало виды внутри рода. В целях идентификации Systema Sexuale Линнея классифицировала растения на 24 группы в зависимости от количества их мужских половых органов. В 24-ю группу, Cryptogamia , вошли все растения со скрытыми репродуктивными органами, мхи, печеночники, папоротники, водоросли и грибы.
Расширение знаний об анатомии , морфологии и жизненных циклах растений привело к осознанию того, что между растениями было больше естественного сходства, чем искусственная половая система Линнея. Адансон (1763 г.), де Жюссье (1789 г.) и Кандоль (1819 г.) предложили различные альтернативные естественные системы классификации, которые группировали растения с использованием более широкого диапазона общих признаков и получили широкое распространение. Система Кандолла отражала его идеи о развитии морфологической сложности, а более поздняя система Бентама и Хукера , которая имела влияние до середины 19 века, находилась под влиянием подхода Кандоля. Публикация Дарвином « Происхождения видов» в 1859 году и его концепции общего происхождения потребовали модификации системы Кандолла, чтобы отразить эволюционные отношения, отличные от простого морфологического сходства.
Ботаника была сильно стимулирована появлением первого «современного» учебника, Grundzüge der Wissenschaftlichen Botanik Маттиаса Шлейдена , опубликованного на английском языке в 1849 году как «Принципы научной ботаники» . Шлейден был микроскопистом и одним из первых анатомов растений, который вместе с Теодором Шванном и Рудольфом Вирховым основал клеточную теорию и был одним из первых, кто осознал значение клеточного ядра , описанного Робертом Брауном в 1831 году. В 1855 году Адольф Фик сформулировал законы Фика, которые позволили рассчитать скорость молекулярной диффузии в биологических системах.

Поздняя современная ботаника

Опираясь на генно-хромосомную теорию наследственности, созданную Грегором Менделем (1822–1884), Август Вейсман (1834–1914) доказал, что наследование происходит только через гаметы . Никакие другие клетки не могут передавать унаследованные признаки. Работа Кэтрин Исау (1898–1997) по анатомии растений до сих пор является важной основой современной ботаники. Ее книги « Анатомия растений» и «Анатомия семенных растений » уже более полувека являются ключевыми учебниками по структурной биологии растений.
Дисциплина экологии растений была впервые введена в конце 19 века такими ботаниками, как Евгений Варминг , выдвинувший гипотезу о том, что растения образуют сообщества , и его наставник и преемник Кристен К. Раункиер , чья система описания форм жизни растений используется до сих пор. Концепция о том, что состав растительных сообществ, таких как широколиственные леса умеренного пояса , изменяется в процессе экологической сукцессии , была разработана Генри Чендлером Коулзом , Артуром Тэнсли и Фредериком Клементсом . Клементсу приписывают идею кульминационной растительности как наиболее сложной растительности, которую может поддерживать окружающая среда, а Тэнсли ввел концепцию экосистем в биологию. Опираясь на обширную более раннюю работу Альфонса де Кандоля , Николай Вавилов (1887–1943) подготовил отчеты о биогеографии , центрах происхождения и истории эволюции хозяйственных растений.
В частности, с середины 1960-х годов произошел прогресс в понимании физики физиологических процессов растений, таких как транспирация (транспорт воды в тканях растений), температурная зависимость скорости испарения воды с поверхности листа и молекулярная диффузия воды. паров и углекислого газа через устьичные отверстия. Эти разработки в сочетании с новыми методами измерения размера устьичных отверстий и скорости фотосинтеза позволили точно описать скорость газообмена между растениями и атмосферой. Инновации в статистическом анализе Рональда Фишера , Фрэнка Йейтса и других на экспериментальной станции Ротамстеда способствовали рациональному планированию экспериментов и анализу данных в ботанических исследованиях. Открытие и идентификация Кеннетом В. Тиманом в 1948 году растительных гормонов ауксинов позволило регулировать рост растений с помощью внешних химических веществ. Фредерик Кэмпион Стюард был пионером методов микроразмножения и культивирования тканей растений, контролируемых растительными гормонами. Синтетический ауксин 2,4-дихлорфеноксиуксусная кислота или 2,4-Д был одним из первых коммерческих синтетических гербицидов.
Развитие биохимии растений в 20 веке было обусловлено современными методами органического химического анализа , такими как спектроскопия , хроматография и электрофорез . С появлением родственных молекулярно-биологических подходов молекулярной биологии , геномики , протеомики и метаболомики взаимосвязь между геномом растения и большинством аспектов биохимии, физиологии, морфологии и поведения растений может быть подвергнута подробному экспериментальному анализу. Концепция, первоначально высказанная Готлибом Хаберландтом в 1902 г., согласно которой все растительные клетки являются тотипотентными и могут быть выращены in vitro , в конечном итоге позволила использовать генную инженерию в эксперименте для выключения гена или генов, ответственных за конкретный признак, или для добавления таких генов, как GFP , которые сообщить , когда экспрессируется интересующий ген. Эти технологии позволяют биотехнологическое использование целых растений или культур растительных клеток, выращенных в биореакторах , для синтеза пестицидов , антибиотиков или других фармацевтических препаратов , а также практическое применение генетически модифицированных сельскохозяйственных культур , предназначенных для повышения урожайности.
Современная морфология признает континуум между основными морфологическими категориями корня, стебля (каулома), листа (филлома) и трихомы . Кроме того, он подчеркивает структурную динамику. Современная систематика стремится отразить и раскрыть филогенетические отношения между растениями. Современная молекулярная филогенетика в значительной степени игнорирует морфологические признаки, полагаясь на последовательности ДНК в качестве данных. Молекулярный анализ последовательностей ДНК большинства семейств цветковых растений позволил Группе филогении покрытосеменных опубликовать в 1998 году филогению цветковых растений, ответив на многие вопросы о взаимоотношениях между семействами и видами покрытосеменных растений. Теоретическая возможность практического метода идентификации видов и коммерческих сортов растений с помощью ДНК-штрихкодирования является предметом активных текущих исследований.
Объем и важность

Изучение растений жизненно важно, потому что они лежат в основе почти всей животной жизни на Земле, производя большую часть кислорода и пищи, которые обеспечивают людей и другие организмы с помощью аэробного дыхания химической энергией, необходимой им для существования. Растения, водоросли и цианобактерии являются основными группами организмов, осуществляющих фотосинтез — процесс, использующий энергию солнечного света для преобразования воды и углекислого газа в сахара, которые можно использовать как в качестве источника химической энергии, так и в качестве источника органических молекул. в структурных компонентах клеток. В качестве побочного продукта фотосинтеза растения выделяют в атмосферу кислород — газ, необходимый почти всем живым существам для осуществления клеточного дыхания. Кроме того, они влияют на глобальные круговороты углерода и воды , а корни растений связывают и стабилизируют почвы, предотвращая эрозию почвы . Растения имеют решающее значение для будущего человеческого общества, поскольку они обеспечивают людей пищей, кислородом, лекарствами и продуктами, а также создают и сохраняют почву.
Исторически все живые существа классифицировались либо как животные, либо как растения, а ботаника охватывала изучение всех организмов, не считавшихся животными. Ботаники исследуют как внутренние функции, так и процессы внутри растительных органелл , клеток, тканей, целых растений, растительных популяций и растительных сообществ. На каждом из этих уровней ботаник может заниматься классификацией ( таксономия ), филогении и эволюции , структурой ( анатомия и морфология ) или функцией ( физиология ) жизни растений.
Самое строгое определение «растение» включает только «наземные растения» или эмбриофиты , к которым относятся семенные растения (голосеменные, в том числе сосны , и цветковые растения ) и свободно спорящие криптогамы , включая папоротники , плауны , печеночники , роголистники и мхи . Эмбриофиты — это многоклеточные эукариоты , произошедшие от предка, который получал энергию от солнечного света путем фотосинтеза . У них есть жизненные циклы с чередованием гаплоидных и диплоидных фаз. Половая гаплоидная фаза эмбриофитов, известная как гаметофит , питает развивающийся спорофит диплоидного зародыша в своих тканях, по крайней мере, часть его жизни, даже в семенных растениях, где сам гаметофит питается своим родительским спорофитом. К другим группам организмов, которые ранее изучались ботаниками, относятся бактерии (сейчас изучаются в бактериологии ), грибы ( микология ), в том числе лихенообразующие грибы ( лихенология ), нехлорофитные водоросли ( фикология ) и вирусы ( вирусология ). Однако ботаники по-прежнему уделяют внимание этим группам, а грибы (в том числе лишайники) и фотосинтезирующие простейшие обычно рассматриваются во вводных курсах ботаники.
Палеоботаники изучают древние растения в летописи окаменелостей, чтобы получить информацию об эволюционной истории растений . Считается, что цианобактерии , первые фотосинтезирующие организмы на Земле, высвобождающие кислород, дали начало предкам растений, вступив в эндосимбиотические отношения с ранними эукариотами, в конечном итоге став хлоропластами в растительных клетках. Новые фотосинтезирующие растения (вместе со своими родственниками-водорослями) ускорили повышение уровня атмосферного кислорода , начатое цианобактериями , изменив древнюю бескислородную, восстановительную атмосферу на такую, в которой свободный кислород был в изобилии более 2 миллиардов лет.
Среди важных ботанических вопросов 21-го века — роль растений как первичных производителей в глобальном круговороте основных компонентов жизни: энергии, углерода, кислорода, азота и воды, а также то, как наша забота о растениях может помочь решить глобальные экологические проблемы управление ресурсами , сохранение , продовольственная безопасность человека , биологически инвазивные организмы , связывание углерода , изменение климата и устойчивость .
Питание человека

Практически все основные продукты питания поступают либо непосредственно из растений, либо косвенно из животных, которые их едят. Растения и другие фотосинтезирующие организмы лежат в основе большинства пищевых цепей, поскольку они используют энергию солнца и питательные вещества из почвы и атмосферы, преобразуя их в форму, пригодную для использования животными. Это то, что экологи называют первым трофическим уровнем . Современные формы основных продуктов питания , таких как конопля , теф , кукуруза, рис, пшеница и другие злаковые травы, бобовые , бананы и плантаны, а также конопля , лен и хлопок , выращенные для их волокон, являются результатом доисторического отбора. на протяжении тысячелетий из числа дикорастущих растений-предков с наиболее желательными характеристиками.
Ботаники изучают, как растения производят пищу и как повысить урожайность, например, путем селекции растений , что делает их работу важной для способности человечества накормить мир и обеспечить продовольственную безопасность для будущих поколений. Ботаники также изучают сорняки, которые представляют значительную проблему в сельском хозяйстве, а также биологию и борьбу с фитопатогенами в сельском хозяйстве и природных экосистемах . Этноботаника – это наука о взаимоотношениях между растениями и людьми. Применительно к исследованию исторических отношений растений и людей этноботанику можно назвать археоботаникой или палеоэтноботаникой . Некоторые из самых ранних отношений растений и людей возникли между коренными жителями Канады при идентификации съедобных растений от несъедобных. Это отношение коренных жителей к растениям было зафиксировано этноботаниками.
Биохимия растений
Биохимия растений - это изучение химических процессов, используемых растениями. Некоторые из этих процессов используются в их первичном метаболизме , например, фотосинтетический цикл Кальвина и метаболизм крассуловой кислоты . Другие производят специальные материалы, такие как целлюлоза и лигнин , используемые для построения их тел, и вторичные продукты, такие как смолы и ароматические соединения .

Растения и различные другие группы фотосинтезирующих эукариот, известных под общим названием « водоросли », имеют уникальные органеллы, известные как хлоропласты . Считается, что хлоропласты произошли от цианобактерий , которые сформировали эндосимбиотические отношения с древними предками растений и водорослей. Хлоропласты и цианобактерии содержат сине-зеленый пигмент хлорофилл а . Хлорофилл а (а также его двоюродный брат хлорофилл b , характерный для растений и зеленых водорослей ) поглощает свет в сине-фиолетовой и оранжево-красной частях спектра , отражая и пропуская зеленый свет, который мы видим как характерный цвет этих организмов. Энергия красного и синего света, которую поглощают эти пигменты, используется хлоропластами для образования богатых энергией углеродных соединений из углекислого газа и воды посредством оксигенного фотосинтеза — процесса, в результате которого в качестве побочного продукта образуется молекулярный кислород (O 2 ).

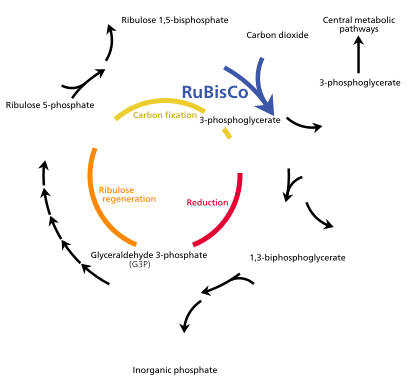


Энергия света, захваченная хлорофиллом а , первоначально находится в форме электронов (а затем протонного градиента ), которые используются для создания молекул АТФ и НАДФН , которые временно хранят и транспортируют энергию. Их энергия используется в светонезависимых реакциях цикла Кальвина ферментом рубиско для производства молекул 3-углеродного сахара глицеральдегид-3-фосфата (G3P). Глицеральдегид-3-фосфат является первым продуктом фотосинтеза и сырьем, из которого синтезируются глюкоза и почти все другие органические молекулы биологического происхождения. Часть глюкозы превращается в крахмал, который запасается в хлоропластах. Крахмал является характерным запасом энергии большинства наземных растений и водорослей, в то время как инулин , полимер фруктозы , используется для той же цели в семействе сложноцветных подсолнечника . Часть глюкозы превращается в сахарозу (обычный столовый сахар) для экспорта на остальную часть завода.
В отличие от животных (у которых отсутствуют хлоропласты), растения и их родственники-эукариоты делегировали своим хлоропластам многие биохимические роли , включая синтез всех своих жирных кислот и большинства аминокислот . Жирные кислоты, которые вырабатывают хлоропласты, используются для многих целей, таких как обеспечение материала для построения клеточных мембран и создание полимера кутина , который содержится в кутикуле растений и защищает наземные растения от высыхания.
Растения синтезируют ряд уникальных полимеров , таких как полисахаридные молекулы целлюлозы , пектин и ксилоглюкан , из которых построена клеточная стенка наземных растений. Сосудистые наземные растения производят лигнин , полимер, используемый для укрепления вторичных клеточных стенок ксилемных трахеид и сосудов , чтобы предотвратить их разрушение, когда растение всасывает через них воду в условиях водного стресса. Лигнин также используется в других типах клеток, таких как волокна склеренхимы , которые обеспечивают структурную поддержку растения и являются основным компонентом древесины. Спорополленин представляет собой химически стойкий полимер, обнаруженный во внешних клеточных стенках спор и пыльцы наземных растений, ответственный за выживание ранних спор наземных растений и пыльцы семенных растений в летописи окаменелостей. Он широко считается маркером начала эволюции наземных растений в ордовикский период. Концентрация углекислого газа в атмосфере сегодня намного ниже, чем была, когда растения появились на суше в ордовикский и силурийский периоды. Многие однодольные , такие как кукуруза и ананас , и некоторые двудольные , такие как сложноцветные , с тех пор независимо развили пути, такие как метаболизм крассуловой кислоты и путь фиксации углерода C 4 для фотосинтеза, которые позволяют избежать потерь в результате фотодыхания в более распространенном пути фиксации углерода C 3 . Эти биохимические стратегии уникальны для наземных растений.
Медицина и материалы
Фитохимия — это раздел биохимии растений, в первую очередь занимающийся химическими веществами, образующимися растениями в ходе вторичного метаболизма . Некоторые из этих соединений являются токсинами, такими как алкалоид кониин болиголова . Другие, такие как эфирные масла перечной мяты и лимонного масла, полезны из-за их аромата, в качестве ароматизаторов и специй (например, капсаицин ), а также в медицине в качестве фармацевтических препаратов, таких как опиум из опийного мака . Многие лекарственные и рекреационные наркотики , такие как тетрагидроканнабинол (активный ингредиент каннабиса ), кофеин , морфин и никотин , получают непосредственно из растений. Другие являются простыми производными ботанических натуральных продуктов. Например, болеутоляющее средство аспирин представляет собой ацетиловый эфир салициловой кислоты , первоначально выделенный из коры ивовых деревьев , а широкий спектр опиатных болеутоляющих средств , таких как героин , получают путем химической модификации морфина , полученного из опийного мака . Популярные стимуляторы получают из растений, таких как кофеин из кофе, чая и шоколада, а никотин из табака. Большинство алкогольных напитков получают путем ферментации богатых углеводами растительных продуктов, таких как ячмень (пиво), рис ( саке ) и виноград (вино). Коренные американцы использовали различные растения для лечения болезней на протяжении тысячелетий. Эти знания коренных американцев о растениях были зафиксированы энтоботаниками , а затем, в свою очередь, использованы фармацевтическими компаниями для открытия новых лекарств .
Растения могут синтезировать полезные цветные красители и пигменты, такие как антоцианы , отвечающие за красный цвет красного вина , желтая сварка и голубая вайда , используемые вместе для производства линкольн-зеленого , индоксил , источник синего красителя индиго , традиционно используемого для окрашивания джинсовой ткани и пигментов для художников. гамбодж и марена розовая . Сахар, крахмал , хлопок, лен , пенька , некоторые виды веревок , древесина и древесностружечные плиты , папирус и бумага, растительные масла , воск и натуральный каучук являются примерами коммерчески важных материалов, изготовленных из растительных тканей или их вторичных продуктов. Древесный уголь , чистая форма углерода, полученная путем пиролиза древесины, имеет долгую историю использования в качестве топлива для плавки металлов, в качестве фильтрующего материала и адсорбента , а также в качестве материала для художников и является одним из трех ингредиентов пороха . Целлюлоза , самый распространенный в мире органический полимер, может быть преобразована в энергию, топливо, материалы и химическое сырье. Изделия из целлюлозы включают вискозу и целлофан , клей для обоев , биобутанол и пушечную вату . Сахарный тростник , рапс и соя являются некоторыми из растений с высоким содержанием сахара или масла, способных к брожению, которые используются в качестве источников биотоплива , важной альтернативы ископаемому топливу , такому как биодизель . Сладкая трава использовалась коренными американцами для защиты от таких насекомых, как комары . Эти отпугивающие насекомых свойства душистой травы были позже обнаружены Американским химическим обществом в молекулах фитола и кумарина .
Экология растений

Экология растений — это наука о функциональных отношениях между растениями и их средами обитания — средами, в которых они завершают свой жизненный цикл . Экологи растений изучают состав местных и региональных флор , их биоразнообразие , генетическое разнообразие и приспособленность , адаптацию растений к окружающей среде и их конкурентные или мутуалистические взаимодействия с другими видами. Некоторые экологи даже полагаются на эмпирические данные коренных народов, собранные этноботаниками. Эта информация может передать большой объем информации о том, какой была земля тысячи лет назад и как она изменилась за это время. Цели экологии растений - понять причины их моделей распространения, продуктивности, воздействия на окружающую среду, эволюции и реакции на изменение окружающей среды.
Растения зависят от определенных эдафических (почвенных) и климатических факторов окружающей среды, но также могут изменять эти факторы. Например, они могут изменять альбедо своей среды , увеличивать перехват стока , стабилизировать минеральные почвы и развивать их органическое содержание, а также влиять на локальную температуру. Растения конкурируют с другими организмами в своей экосистеме за ресурсы. Они взаимодействуют со своими соседями в различных пространственных масштабах в группах, популяциях и сообществах , которые в совокупности составляют растительность. Регионы с характерными типами растительности и доминирующими растениями, а также с аналогичными абиотическими и биотическими факторами, климатом и географией составляют биомы , такие как тундра или тропический лес .
Травоядные едят растения, но растения могут защищать себя, а некоторые виды паразитируют или даже плотоядны . Другие организмы образуют взаимовыгодные отношения с растениями. Например, микоризные грибы и ризобии обеспечивают растения питательными веществами в обмен на пищу, муравьиные растения привлекают муравьиных растений для защиты, медоносные пчелы , летучие мыши и другие животные опыляют цветы, а люди и другие животные действуют как переносчики для распространения спор и семян .
Растения, климат и изменение окружающей среды
Реакция растений на климат и другие изменения окружающей среды может помочь нам понять, как эти изменения влияют на функционирование и продуктивность экосистемы. Например, фенология растений может быть полезным косвенным показателем температуры в исторической климатологии и биологического воздействия изменения климата и глобального потепления . Палинология , анализ отложений ископаемой пыльцы в отложениях тысяч или миллионов лет назад , позволяет реконструировать климат прошлого. Оценки концентраций CO 2 в атмосфере с палеозоя были получены на основании плотности устьиц , а также формы и размеров листьев древних наземных растений . Истощение озонового слоя может подвергнуть растения воздействию более высоких уровней ультрафиолетового излучения-B (UV-B), что приведет к снижению скорости роста. Более того, информация, полученная в результате исследований в области экологии сообществ , систематики растений и таксономии , необходима для понимания изменений растительности , разрушения среды обитания и исчезновения видов .
Генетика

Наследование у растений следует тем же фундаментальным принципам генетики, что и у других многоклеточных организмов. Грегор Мендель открыл генетические законы наследования , изучая унаследованные черты, такие как форма у Pisum sativum ( горох ). То, что Мендель узнал из изучения растений, имело далеко идущие преимущества за пределами ботаники. Точно так же « прыгающие гены » были обнаружены Барбарой МакКлинток , когда она изучала кукурузу. Тем не менее, существуют некоторые отличительные генетические различия между растениями и другими организмами.
Границы видов у растений могут быть слабее, чем у животных, и часто возможны межвидовые гибриды . Знакомым примером является мята перечная , Mentha × piperita , стерильный гибрид Mentha aquatica и мяты колосовой, Mentha spicata . Многие культурные сорта пшеницы являются результатом многократных меж- и внутривидовых скрещиваний между дикими видами и их гибридами. Покрытосеменные с однодомными цветками часто имеют механизмы самонесовместимости, которые действуют между пыльцой и рыльцем , так что пыльца либо не достигает рыльца, либо не может прорасти и произвести мужские гаметы . Это один из нескольких методов, используемых растениями для стимулирования ауткроссинга . У многих наземных растений мужские и женские гаметы образуются отдельными особями. Эти виды называются двудомными , когда речь идет о спорофитах сосудистых растений, и двудомными, когда речь идет о гаметофитах мохообразных .
В отличие от высших животных, у которых партеногенез встречается редко, бесполое размножение у растений может происходить по нескольким различным механизмам. Одним из примеров является образование стеблевых клубней у картофеля. В частности, в арктических или альпийских местообитаниях, где возможности оплодотворения цветов животными редки, вместо цветов могут развиваться проростки или луковицы , заменяя половое размножение бесполым и давая начало клональным популяциям , генетически идентичным родительским. Это один из нескольких типов апомиксиса , встречающихся у растений. Апомиксис также может произойти в семени , производя семя, содержащее зародыш, генетически идентичный родителю.
Большинство организмов, размножающихся половым путем , являются диплоидными, с парными хромосомами, но удвоение числа их хромосом может происходить из-за ошибок в цитокинезе . Это может произойти на ранней стадии развития с образованием аутополиплоидного или частично аутополиплоидного организма, или во время нормальных процессов клеточной дифференцировки с образованием некоторых типов полиплоидных клеток ( эндополиплоидия ), или во время образования гамет . Аллополиплоидное растение может возникнуть в результате гибридизации между двумя разными видами. И автополиплоидные, и аллополиплоидные растения часто могут нормально воспроизводиться, но не могут успешно скрещиваться с родительской популяцией из-за несоответствия числа хромосом. Эти растения, которые репродуктивно изолированы от родительских видов, но живут в пределах одного и того же географического района, могут быть достаточно успешными для образования нового вида . Некоторые стерильные полиплоиды растений все еще могут воспроизводиться вегетативно или путем апомиксиса семян, образуя клональные популяции идентичных особей. Твердые сорта пшеницы являются фертильными тетраплоидными аллополиплоидами, а мягкая пшеница — фертильными гексаплоидами . Коммерческий банан является примером стерильного триплоидного гибрида без косточек. Одуванчик обыкновенный - триплоид, дающий жизнеспособные семена за счет апомиктических семян.
Как и у других эукариот, наследование эндосимбиотических органелл, таких как митохондрии и хлоропласты , у растений неменделевское . Хлоропласты наследуются от мужского родителя у голосеменных, но часто от женского родителя у цветковых растений.
Молекулярная генетика

Значительный объем новых знаний о функциях растений получен в результате исследований молекулярной генетики модельных растений , таких как кресс-салат Тале, Arabidopsis thaliana , сорных видов семейства горчичных ( Brassicaceae ). Геном или наследственная информация, содержащаяся в генах этого вида, кодируется примерно 135 миллионами пар оснований ДНК, образуя один из самых маленьких геномов среди цветковых растений . Arabidopsis был первым растением, геном которого был секвенирован в 2000 году. Секвенирование некоторых других относительно небольших геномов, риса ( Oryza sativa ) и Brachypodium distachyon , сделало их важными модельными видами для понимания генетики, клеточной и молекулярной биологии злаков . , травы и однодольные в целом.
Модельные растения , такие как Arabidopsis thaliana , используются для изучения молекулярной биологии растительных клеток и хлоропластов . В идеале эти организмы имеют небольшие геномы, которые хорошо известны или полностью секвенированы, небольшой рост и короткое время генерации. Кукуруза использовалась для изучения механизмов фотосинтеза и насыщения флоэмы сахаром растений C 4 . Одноклеточная зеленая водоросль Chlamydomonas reinhardtii , хотя сама по себе и не является эмбриофитом , содержит хлоропласт с зеленым пигментом, сходный с хлоропластом наземных растений, что делает ее полезной для изучения. Красная водоросль Cyanidioschyzon merolae также использовалась для изучения некоторых основных функций хлоропластов. Шпинат , горох , соевые бобы и мох Physcomitrella patens обычно используются для изучения биологии клеток растений.
Agrobacterium tumefaciens , почвенная ризосферная бактерия, может прикрепляться к растительным клеткам и заражать их каллус -индуцирующей плазмидой Ti путем горизонтального переноса генов , вызывая каллусную инфекцию, называемую болезнью корончатого галла. Шелл и Ван Монтегю (1977) предположили, что плазмида Ti может быть естественным вектором для введения гена Nif, ответственного за фиксацию азота , в клубеньках бобовых и других видов растений. На сегодняшний день генетическая модификация плазмиды Ti является одним из основных методов введения трансгенов в растения и создания генетически модифицированных культур .
Эпигенетика
Эпигенетика - это изучение наследственных изменений в функции генов , которые нельзя объяснить изменениями в основной последовательности ДНК, но которые заставляют гены организма вести себя (или «выражать себя») по-разному. Одним из примеров эпигенетических изменений является маркировка генов путем метилирования ДНК , которая определяет, будут ли они экспрессироваться или нет. Экспрессия генов также может контролироваться белками-репрессорами, которые прикрепляются к сайленсерным областям ДНК и предотвращают экспрессию этой области кода ДНК. Эпигенетические метки могут быть добавлены или удалены из ДНК на запрограммированных стадиях развития растения и ответственны, например, за различия между пыльниками, лепестками и нормальными листьями, несмотря на то, что все они имеют один и тот же основной генетический код. Эпигенетические изменения могут быть временными или могут сохраняться в результате последовательных клеточных делений до конца жизни клетки. Было показано, что некоторые эпигенетические изменения передаются по наследству , в то время как другие восстанавливаются в зародышевых клетках.
Эпигенетические изменения в биологии эукариот служат для регуляции процесса клеточной дифференцировки . Во время морфогенеза тотипотентные стволовые клетки становятся различными плюрипотентными клеточными линиями эмбриона , которые , в свою очередь, становятся полностью дифференцированными клетками. Одна оплодотворенная яйцеклетка, зигота , дает начало многим различным типам растительных клеток , включая паренхиму , элементы сосудов ксилемы , ситовидные трубки флоэмы , замыкающие клетки эпидермиса и т. д ., поскольку она продолжает делиться . Этот процесс является результатом эпигенетической активации одних генов и ингибирования других.
В отличие от животных, многие растительные клетки, особенно клетки паренхимы , окончательно не дифференцируются, оставаясь тотипотентными со способностью давать начало новому индивидуальному растению. Исключения составляют сильно одревесневшие клетки, склеренхима и ксилема, которые отмирают при созревании, и ситовидные трубки флоэмы без ядер. В то время как растения используют многие из тех же эпигенетических механизмов, что и животные, такие как ремоделирование хроматина , альтернативная гипотеза состоит в том, что растения устанавливают свои паттерны экспрессии генов, используя информацию о положении из окружающей среды и окружающих клеток, чтобы определить судьбу своего развития.
Эпигенетические изменения могут привести к парамутациям , которые не подчиняются правилам менделевского наследия. Эти эпигенетические метки передаются от одного поколения к другому, причем один аллель вызывает изменение другого.
Эволюция растений

Хлоропласты растений имеют ряд биохимических, структурных и генетических сходств с цианобактериями (обычно, но неправильно называемыми «сине-зелеными водорослями») и, как полагают, произошли от древних эндосимбиотических отношений между предковой эукариотической клеткой и цианобактериальной резидентной клеткой. .
Водоросли представляют собой полифилетическую группу и относятся к различным отделам, некоторые из которых более тесно связаны с растениями, чем другие. Между ними есть много различий в таких особенностях, как состав клеточной стенки, биохимия, пигментация, структура хлоропластов и запасы питательных веществ. Подразделение водорослей Charophyta , сестра отдела зеленых водорослей Chlorophyta , считается предком настоящих растений. Класс Charophyte Charophyceae и подцарство наземных растений Embryophyta вместе образуют монофилетическую группу или кладу Streptophytina .
Несосудистые наземные растения — эмбриофиты , у которых отсутствуют ксилема и флоэма сосудистых тканей . К ним относятся мхи , печеночники и роголистники . Птеридофитные сосудистые растения с настоящей ксилемой и флоэмой, которые размножались спорами, прорастающими в свободноживущие гаметофиты, эволюционировали в силурийский период и диверсифицировались в несколько линий в течение позднего силура и раннего девона . Представители плауновидных сохранились до наших дней. К концу девонского периода несколько групп, в том числе плауновидные , сфенофиллы и прогимноспермы , независимо развили «мегаспоры» — их споры были двух разных размеров: более крупные мегаспоры и более мелкие микроспоры. Их уменьшенные гаметофиты развились из мегаспор, оставшихся в спорообразующих органах (мегаспорангиях) спорофита, состояние, известное как эндоспория. Семена состоят из эндоспорного мегаспорангия, окруженного одним или двумя покровными слоями ( интегументами ). Молодой спорофит развивается внутри семени, которое при прорастании расщепляется, чтобы высвободить его. Самые ранние известные семенные растения относятся к позднему девонскому фаменскому этапу. После эволюции семенного габитуса семенные растения диверсифицировались, что привело к появлению ряда ныне вымерших групп, включая семенные папоротники , а также современные голосеменные и покрытосеменные растения . Голосеменные растения дают «голые семена», не полностью заключенные в завязь; к современным представителям относятся хвойные деревья , саговники , гинкго , гнеталес . Покрытосеменные производят семена, заключенные в структуру, такую как плодолистик или завязь . Текущие исследования молекулярной филогенетики живых растений, по-видимому, показывают, что покрытосеменные растения являются родственными ветвями голосеменных растений.
Физиология растений

Физиология растений охватывает всю внутреннюю химическую и физическую активность растений, связанную с жизнью. Химические вещества, получаемые из воздуха, почвы и воды, составляют основу всего метаболизма растений . Энергия солнечного света, улавливаемая оксигенным фотосинтезом и высвобождаемая клеточным дыханием , является основой почти всей жизни. Фотоавтотрофы , в том числе все зеленые растения, водоросли и цианобактерии , получают энергию непосредственно от солнечного света путем фотосинтеза. Гетеротрофы , включая всех животных, все грибы, все полностью паразитические растения и нефотосинтезирующие бактерии, поглощают органические молекулы, произведенные фотоавтотрофами, и вдыхают их или используют для построения клеток и тканей. Дыхание — это окисление соединений углерода путем их расщепления на более простые структуры с высвобождением содержащейся в них энергии, что по существу противоположно фотосинтезу.
Молекулы перемещаются внутри растений в результате транспортных процессов, действующих в различных пространственных масштабах . Субклеточный транспорт ионов, электронов и молекул, таких как вода и ферменты, происходит через клеточные мембраны . Минеральные вещества и вода переносятся от корней к другим частям растения с потоком транспирации . Диффузия , осмос , активный транспорт и массовый поток — все это различные способы переноса. Примерами элементов, которые растениям необходимо транспортировать, являются азот , фосфор , калий , кальций , магний и сера . У сосудистых растений эти элементы извлекаются из почвы в виде растворимых ионов корнями и транспортируются по растению в ксилеме. Большинство элементов, необходимых для питания растений , получают в результате химического распада почвенных минералов. Сахароза , полученная в результате фотосинтеза, транспортируется из листьев в другие части растения по флоэме, а растительные гормоны транспортируются с помощью различных процессов.
Гормоны растений

2 Когда солнце расположено под углом и светит только на одну сторону побега, ауксин перемещается на противоположную сторону и стимулирует там удлинение клеток .
3 и 4 Дополнительный рост на этой стороне заставляет побег наклоняться к солнцу .
Растения не пассивны, а реагируют на внешние сигналы , такие как свет, прикосновение и травмы, перемещаясь или увеличиваясь в направлении или в направлении от раздражителя, в зависимости от ситуации. Вещественным свидетельством тактильной чувствительности является почти мгновенное схлопывание листочков Mimosa pudica , ловушек для насекомых венериной мухоловки и пузырчатки , а также поллиний орхидей.
Гипотеза о том, что рост и развитие растений координируется растительными гормонами или регуляторами роста растений, впервые возникла в конце 19 века. Дарвин экспериментировал с движением побегов и корней растений навстречу свету и гравитации и пришел к выводу: «Вряд ли будет преувеличением сказать, что кончик корешка… действует как мозг одного из низших животных… направляя несколько движений». . Примерно в то же время роль ауксинов (от греч . auxein — расти) в контроле роста растений впервые обозначила голландский ученый Фриц Вент . Первый известный ауксин, индол-3-уксусная кислота (ИУК), который способствует росту клеток, был выделен из растений только примерно 50 лет спустя. Это соединение опосредует тропические реакции побегов и корней на свет и гравитацию. Обнаружение в 1939 г. того, что каллус растений можно поддерживать в культуре, содержащей ИУК, а затем наблюдение в 1947 г., что его можно стимулировать к образованию корней и побегов путем контроля концентрации гормонов роста, были ключевыми шагами в развитии биотехнологии растений и генетической модификации. .
Цитокинины представляют собой класс растительных гормонов, названных в честь их контроля деления клеток (особенно цитокинеза ). Природный цитокинин зеатин был обнаружен в кукурузе Zea mays и является производным пурина аденина . Зеатин вырабатывается в корнях и транспортируется к побегам в ксилеме, где он способствует делению клеток, развитию почек и озеленению хлоропластов. Гибберелины , такие как гибберелиновая кислота , представляют собой дитерпены , синтезируемые из ацетил-КоА мевалонатным путем . Они участвуют в содействии прорастанию и нарушению покоя семян, в регулировании высоты растений путем контроля удлинения стебля и контроля цветения. Абсцизовая кислота (АБК) встречается во всех наземных растениях, кроме печеночников, и синтезируется из каротиноидов в хлоропластах и других пластидах. Он ингибирует деление клеток, способствует созреванию семян и переходу в состояние покоя, а также способствует закрытию устьиц. Он был назван так потому, что изначально считалось, что он контролирует опадение . Этилен — газообразный гормон, образующийся во всех тканях высших растений из метионина . В настоящее время известно, что это гормон, который стимулирует или регулирует созревание и опадение плодов, и он, или синтетический регулятор роста этефон , который быстро метаболизируется с образованием этилена, используется в промышленных масштабах для ускорения созревания хлопка, ананасов и других климактерических культур . .
Другой класс фитогормонов - это жасмонаты , впервые выделенные из масла Jasminum grandiflorum , которые регулируют реакцию растений на рану, разблокируя экспрессию генов, необходимых для системной приобретенной реакции устойчивости к атаке патогенов.
Помимо того, что свет является основным источником энергии для растений, он действует как сигнальное устройство, предоставляя растениям информацию, например, о том, сколько солнечного света растение получает каждый день. Это может привести к адаптивным изменениям в процессе, известном как фотоморфогенез . Фитохромы — фоторецепторы растений, чувствительные к свету.
Анатомия и морфология растений

Анатомия растений — это изучение строения растительных клеток и тканей, тогда как морфология растений — это изучение их внешней формы. Все растения — многоклеточные эукариоты, их ДНК хранится в ядрах. Характерные особенности растительных клеток , которые отличают их от клеток животных и грибов, включают первичную клеточную стенку , состоящую из полисахаридов целлюлозы , гемицеллюлозы и пектина , более крупные вакуоли , чем в клетках животных, и наличие пластид с уникальными фотосинтетическими и биосинтетическими функциями, как в клетках животных. хлоропласты. Другие пластиды содержат продукты хранения, такие как крахмал ( амилопласты ) или липиды ( элайопласты ). Уникально то, что клетки стрептофитов и зеленых водорослей отряда Trentepohliales делятся путем построения фрагмопласта в качестве шаблона для построения клеточной пластинки в конце клеточного деления .
![Схема «типичного» эвдикота, самого распространенного вида растений (три пятых всех видов растений)[178]. Однако на самом деле ни одно растение не выглядит точно так же.](https://wikipedia.org/wiki/File:Plant.svg)
Тела сосудистых растений , включая плауны , папоротники и семенные растения ( голосеменные и покрытосеменные ), обычно имеют надземную и подземную подсистемы. Побеги состоят из стеблей , несущих зеленые фотосинтезирующие листья и репродуктивные структуры. Подземные васкуляризированные корни несут корневые волоски на концах и обычно лишены хлорофилла. Несосудистые растения, печеночники , роголистники и мхи не образуют проникающих сквозь землю сосудистых корней, и большая часть растений участвует в фотосинтезе. Генерация спорофитов не является фотосинтезирующей у печеночников, но может обеспечивать часть своих энергетических потребностей за счет фотосинтеза у мхов и роголистников.
Корневая система и система побегов взаимозависимы - обычно нефотосинтезирующая корневая система зависит от системы побегов для питания, а обычно фотосинтезирующая система побегов зависит от воды и минералов из корневой системы. Клетки каждой системы способны создавать клетки другой и производить придаточные побеги или корни. Столоны и клубни являются примерами побегов, которые могут укореняться. Корни, которые распространяются близко к поверхности, например, у ивы, могут дать побеги и, в конечном итоге, новые растения. В случае утраты одной из систем другая часто может отрастить ее заново. На самом деле можно вырастить целое растение из одного листа, как в случае с растениями Streptocarpus sect. Сенполия или даже отдельная клетка , которая может дедифференцироваться в каллюс (массу неспециализированных клеток), из которого может вырасти новое растение. У сосудистых растений ксилема и флоэма являются проводящими тканями, которые транспортируют ресурсы между побегами и корнями. Корни часто приспособлены для хранения пищевых продуктов, таких как сахар или крахмал , как у сахарной свеклы и моркови.
Стебли в основном обеспечивают поддержку листьев и репродуктивных структур, но могут хранить воду в суккулентных растениях, таких как кактусы , пищу, как в клубнях картофеля , или размножаться вегетативно , как в столонах растений земляники , или в процессе отводков . Листья собирают солнечный свет и осуществляют фотосинтез . Большие, плоские, гибкие, зеленые листья называются лиственными. Голосеменные , такие как хвойные , саговники , гинкго и гнетофиты , являются семенными растениями с открытыми семенами. Покрытосеменные - это семенные растения , которые производят цветы и имеют заключенные семена. Древесные растения, такие как азалии и дубы , проходят вторичную фазу роста, в результате которой образуются два дополнительных типа тканей: древесина (вторичная ксилема ) и кора (вторичная флоэма и пробка ). Все голосеменные и многие покрытосеменные — древесные растения. Некоторые растения размножаются половым путем, некоторые бесполым, а некоторые обоими способами.
Хотя ссылки на основные морфологические категории, такие как корень, стебель, лист и трихом, полезны, следует помнить, что эти категории связаны через промежуточные формы, так что получается континуум между категориями. Более того, структуры можно рассматривать как процессы, то есть комбинации процессов.
Систематическая ботаника

Систематическая ботаника является частью систематической биологии, которая занимается диапазоном и разнообразием организмов и их взаимоотношений, особенно в том, что определяется их эволюционной историей. Он включает или связан с биологической классификацией, научной таксономией и филогенетикой . Биологическая классификация — это метод, с помощью которого ботаники группируют организмы в такие категории, как роды или виды . Биологическая классификация является формой научной таксономии . Современная таксономия уходит корнями в работу Карла Линнея , который сгруппировал виды в соответствии с общими физическими характеристиками. С тех пор эти группировки были пересмотрены, чтобы лучше соответствовать дарвиновскому принципу общего происхождения - группировка организмов по происхождению, а не по поверхностным характеристикам . В то время как ученые не всегда согласны с тем, как классифицировать организмы, молекулярная филогенетика , которая использует последовательности ДНК в качестве данных, привела к многочисленным недавним пересмотрам эволюционных линий и, вероятно, продолжит это делать. Господствующая система классификации называется линнеевской таксономией . Она включает в себя ранги и биномиальную номенклатуру . Номенклатура ботанических организмов кодифицирована в Международном кодексе номенклатуры водорослей, грибов и растений (ICN) и управляется Международным ботаническим конгрессом .
Королевство Plantae принадлежит домену Eukarya и рекурсивно разбивается до тех пор, пока каждый вид не будет классифицирован отдельно. Порядок такой: Королевство ; Тип (или Отдел); Класс ; Заказ ; Семья ; Род (множественное число родов ); Виды . Научное название растения представляет его род и его виды в пределах рода, что приводит к единому всемирному названию для каждого организма. Например, тигровая лилия — Lilium columbianum . Lilium — это род, а columbianum — видовой эпитет . Комбинация является названием вида. При написании научного названия организма уместно писать первую букву в роде с заглавной буквы, а все видовые эпитеты — строчными. Кроме того, весь термин обычно выделен курсивом (или подчеркнут, если курсив недоступен).
Эволюционные связи и наследственность группы организмов называют её филогенезом . Филогенетические исследования пытаются открыть филогении. Основной подход заключается в использовании подобия, основанного на общем наследовании, для определения отношений. Например, виды Pereskia представляют собой деревья или кустарники с выступающими листьями. Они явно не напоминают типичный безлистный кактус , такой как эхинокактус . Однако и у Pereskia , и у Echinocactus есть шипы, образованные из ареол (узкоспециализированных подушечек), что позволяет предположить, что эти два рода действительно связаны между собой.


Оценка отношений на основе общих признаков требует осторожности, поскольку растения могут быть похожи друг на друга в результате конвергентной эволюции , в которой признаки возникли независимо. Некоторые молочайные имеют безлистные округлые тела, приспособленные к сохранению воды, как у шаровидных кактусов, но такие признаки, как структура их цветков, ясно показывают, что эти две группы не имеют близкого родства. Кладистический метод использует систематический подход к признакам, различая признаки, не несущие информации об общей эволюционной истории, например, развившиеся отдельно в разных группах ( гомоплазии ) или оставшиеся от предков ( плезиоморфии ), и производные признаки, которые были передаются от нововведений к общему предку ( апоморфии ). Только производные признаки, такие как образующие шипы ареолы кактусов, свидетельствуют о происхождении от общего предка. Результаты кладистического анализа выражаются в виде кладограмм : древовидных диаграмм, показывающих модель эволюционного ветвления и происхождения.
Начиная с 1990-х годов преобладающим подходом к построению филогении живых растений была молекулярная филогенетика , которая использует молекулярные признаки, особенно последовательности ДНК , а не морфологические признаки, такие как наличие или отсутствие шипов и ареол. Разница в том, что сам генетический код используется для определения эволюционных отношений, а не косвенно через признаки, которые он порождает. Клайв Стейс описывает это как наличие «прямого доступа к генетической основе эволюции». В качестве простого примера, до использования генетических данных грибы считались либо растениями, либо более тесно связаны с растениями, чем с животными. Генетические данные свидетельствуют о том, что истинное эволюционное родство многоклеточных организмов показано на кладограмме ниже — грибы более тесно связаны с животными, чем с растениями.
|
|||||||||||||
В 1998 году Группа филогении покрытосеменных опубликовала филогению цветковых растений, основанную на анализе последовательностей ДНК большинства семейств цветковых растений. В результате этой работы были даны ответы на многие вопросы, например, какие семейства представляют самые ранние ветви покрытосеменных растений . Изучение того, как виды растений связаны друг с другом, позволяет ботаникам лучше понять процесс эволюции растений. Несмотря на изучение модельных растений и все более широкое использование доказательств ДНК, среди систематиков продолжается работа и обсуждение того, как лучше всего классифицировать растения по различным таксонам . Технологические разработки, такие как компьютеры и электронные микроскопы , значительно повысили уровень детализации и скорость анализа данных.
Символы
Несколько символов в настоящее время используются в ботанике. Ряд других устарел; например, Линней использовал планетарные символы для древесных, травянистых и многолетних растений, а Уиллд использовал ♄ (Сатурн) для среднего рода в дополнение к ☿ (Меркурий) для обозначения гермафродитов. Следующие символы все еще используются:
- ♀ женщина
- ♂ самец
- ⚥ гермафродит/бисексуал
- ⚲ вегетативное (бесполое) размножение
- ◊ пол неизвестен
- ☉ годовой
-
 (⚇) раз в два года
(⚇) раз в два года
-
 (♾) многолетник
(♾) многолетник
- ☠ ядовитый
- 🛈 дополнительная информация
- × помесный гибрид
- + привитой гибрид
Смотрите также
- Отрасли ботаники
- Эволюция растений
- Глоссарий ботанических терминов
- Глоссарий морфологии растений
- Список журналов по ботанике
- Список ботаников
- Список ботанических садов
- Список ботаников по аббревиатуре автора
- Список одомашненных растений
- Список цветов
- Список систем систематики растений
- Очерк ботаники
- Хронология британской ботаники
Заметки
использованная литература
Цитаты
Источники
- Ачарья, Дипак; Аншу, Шривастава (2008). Местные травяные лекарства: племенные составы и традиционные травяные практики . Джайпур, Индия: Издательство Аавишкар. ISBN 978-81-7910-252-7.
- Аддельсон, Барбара (декабрь 2003 г.). «Естественно-научный институт ботаники и экологии для учителей начальных классов» . Международная организация по охране ботанических садов. Архивировано из оригинала 23 мая 2013 года . Проверено 8 июня 2013 г. .
- Андерсон, Эдвард Ф. (2001). Семья кактусов . Пентленд, Орегон: Timber Press. ISBN 978-0-88192-498-5.
- Армстронг, Джорджия; Херст, Дж. Э. (1996). «Каротиноиды 2: генетика и молекулярная биология биосинтеза каротиноидного пигмента». ФАСЭБ Дж . 10 (2): 228–237. doi : 10.1096/fasebj.10.2.8641556 . PMID 8641556 . S2CID 22385652 .
- Беккер, Буркхард; Марин, Биргер (2009). «Стрептофитные водоросли и происхождение эмбриофитов» . Анналы ботаники . 103 (7): 999–1004. doi : 10.1093/aob/mcp044 . ПВК 2707909 . PMID 19273476 .
- Берлинг, ди-джей ; Осборн, CP; Чалонер, В.Г. (2001). «Эволюция формы листьев у наземных растений, связанная со снижением содержания CO2 в атмосфере в позднепалеозойскую эру» (PDF) . Природа . 410 (6826): 352–354. Бибкод : 2001Natur.410..352B . дои : 10.1038/35066546 . PMID 11268207 . S2CID 4386118 .
- Бендерот, Маркус; Текстор, Сюзанна; Виндзор, Аарон Дж.; Митчелл-Олд, Томас; Гершензон, Джонатан; Кройманн, Юрген (июнь 2006 г.). «Позитивный отбор, стимулирующий диверсификацию вторичного метаболизма растений» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (24): 9118–9123. Бибкод : 2006PNAS..103.9118B . doi : 10.1073/pnas.0601738103 . JSTOR 30051907 . ПВК 1482576 . PMID 16754868 .
- Бен-Менахем, Ари (2009). Историческая энциклопедия естественных и математических наук . Том. 1. Берлин: Springer-Verlag. ISBN 978-3-540-68831-0.
- Беннетт, Чарльз Э .; Хаммонд, Уильям А. (1902). Персонажи Теофраста - Введение . Лондон: Лонгманс, Грин и Ко . Проверено 27 июня 2012 г.
- Беннетт, К.Д.; Уиллис, К.Дж. (2001). "Цветочная пыльца". В Смоле, Джон П.; Биркс, Х. Джон Б. (ред.). Отслеживание изменений окружающей среды с помощью озерных отложений . Том. 3: Наземные, водорослевые и кремнистые индикаторы. Дордрехт, Германия: Kluwer Academic Publishers.
- Берд, Адриан (май 2007 г.). «Восприятие эпигенетики» . Природа . 447 (7143): 396–398. Бибкод : 2007Natur.447..396B . doi : 10.1038/nature05913 . PMID 17522671 . S2CID 4357965 .
- Бьёрн, LO; Каллаган, ТВ; Герке, К.; Йохансон, У .; Сонессон, М. (ноябрь 1999 г.). «Истощение озонового слоя, ультрафиолетовое излучение и жизнь растений». Хемосфера – наука о глобальных изменениях . 1 (4): 449–454. Бибкод : 1999ChGCS...1..449B . doi : 10.1016/S1465-9972(99)00038-0 .
- Жирный, ХК (1977). Царство растений (4-е изд.). Энглвуд Клиффс, Нью-Джерси: Прентис-Холл. ISBN 978-0-13-680389-8.
- Бразелтон, JP (2013). «Что такое биология растений?» . Университет Огайо. Архивировано из оригинала 24 сентября 2015 года . Проверено 3 июня 2013 г. .
- Бургер, Уильям С. (2013). «Происхождение покрытосеменных: первый сценарий однодольных» . Чикаго: Полевой музей.
- Берроуз, WJ (1990). Процессы смены растительности . Лондон: Анвин Хайман. ISBN 978-0-04-580013-1.
- Буц, Стивен Д. (2007). Наука о земных системах (2-е изд.). Клифтон-Парк, Нью-Йорк: Обучение Delmar Cengage. ISBN 978-1-4180-4122-9.
- Кэмпбелл, Нил А .; Рис, Джейн Б.; Урри, Лиза Андреа; Каин, Майкл Л.; Вассерман, Стивен Александр; Минорский, Петр В .; Джексон, Роберт Брэдли (2008). Биология (8-е изд.). Сан-Франциско: Пирсон – Бенджамин Каммингс. ISBN 978-0-321-54325-7.
- де Кандоль, Альфонс (2006). Происхождение культурных растений . Национальный парк Глейшер, Монтана: Kessinger Publishing. ISBN 978-1-4286-0946-4.
- Капон, Брайан (2005). Ботаника для садоводов (2-е изд.). Портленд, Орегон: Timber Publishing. ISBN 978-0-88192-655-2.
- Кавалье-Смит, Томас (2004). «Только шесть королевств жизни» (PDF) . Труды Лондонского королевского общества B . 271 (1545): 1251–1262. doi : 10.1098/rspb.2004.2705 . ПВК 1691724 . PMID 15306349 .
- Чаффи, Найджел (2007). «Анатомия растений, меристемы, клетки и ткани растительного тела Исава: их структура, функции и развитие» . Анналы ботаники . 99 (4): 785–786. doi : 10.1093/aob/mcm015 . ПВК 2802946 .
- Чепмен, Жасмин; Хорсфолл, Питер; О'Брайен, Пэт; Мерфи, Ян; Макдональд, Аверил (2001). Научная сеть . Челтнем, Великобритания: Нельсон Торнс. ISBN 978-0-17-438746-6.
- Чейз, Марк В.; Бремер, Биргитта; Бремер, Коре; Выявить, Джеймс Л.; Солтис, Дуглас Э.; Солтис, Памела С .; Стивенс, Питер С. (2003). «Обновление классификации филогенных групп покрытосеменных для отрядов и семейств цветковых растений: APG II» (PDF) . Ботанический журнал Линнеевского общества . 141 (4): 399–436. doi : 10.1046/j.1095-8339.2003.t01-1-00158.x .
- Чини, А .; Фонсека, С.; Фернандес, Г.; Ади, Б.; Чико, Дж. М.; Лоренцо, О .; Гарсия-Касадо, Г.; Лопес-Видриеро, И.; Лосано, FM; Понсе, МР; Миколь, Дж. Л.; Солано, Р. (2007). «Семейство репрессоров JAZ - недостающее звено в передаче сигналов жасмоната». Природа . 448 (7154): 666–671. Бибкод : 2007Natur.448..666C . doi : 10.1038/nature06006 . PMID 17637675 . S2CID 4383741 .
- Кокинг, Эдвард С. (18 октября 1993 г.). «Некролог: профессор Ф.К. Стюард» . Независимый . Лондон . Проверено 5 июля 2013 г. .
- Коупленд, Герберт Фолкнер (1938). «Царства организмов». Ежеквартальный обзор биологии . 13 (4): 383–420. дои : 10.1086/394568 . S2CID 84634277 .
- Коста, Сильвия; Шоу, Питер (март 2007 г.). "«Открытые» клетки: как клетки могут изменить судьбу » (PDF) . Тенденции в клеточной биологии . 17 (3): 101–106. doi : 10.1016/j.tcb.2006.12.005 . PMID 17194589. Архивировано из оригинала ( PDF)) от 15.12.2013.
- Казенс, Роджер; Мортимер, Мартин (1995). Динамика численности сорняков . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-49969-9.
- Даллал, Ахмад (2010). Ислам, наука и вызов истории . Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 978-0-300-15911-0.
- Дарвин, Чарльз (1880 г.). Сила движения растений (PDF) . Лондон: Мюррей.
- Демоле, Э.; Ледерер, Э.; Мерсье, Д. (1962). «Изоляция и определение структуры жасмоната метила, составляющая характерного запаха эссенции жасмина, изоляция и определение структуры жасмоната метила, составляющая характерного запаха эссенции жасмина». Гельветика Химика Акта . 45 (2): 675–685. doi : 10.1002/hlca.19620450233 .
- Девос, Кэтрин М. ; Гейл, доктор медицины (май 2000 г.). «Отношения генома: модель травы в текущих исследованиях» . Растительная клетка . 12 (5): 637–646. дои : 10.2307/3870991 . JSTOR 3870991 . ПВК 139917 . PMID 10810140 .
- Эрхардт, Д. У.; Фроммер, WB (февраль 2012 г.). «Новые технологии для науки о растениях 21 века» . Растительная клетка . 24 (2): 374–394. doi : 10.1105/tpc.111.093302 . ПВК 3315222 . PMID 22366161 .
- Эрешевски, Марк (1997). «Эволюция линнеевской иерархии». Биология и философия . 12 (4): 493–519. дои : 10.1023/A:1006556627052 . S2CID 83251018 .
- Ферро, Мириам; Сальви, Даниэль; Ривьер-Роллан, Элен; Верма, Тьерри; и другие. (20 августа 2002 г.). «Интегральные мембранные белки оболочки хлоропласта: идентификация и субклеточная локализация новых транспортеров» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (17): 11487–11492. Бибкод : 2002PNAS...9911487F . doi : 10.1073/pnas.172390399 . ПМС 123283 . PMID 12177442 .
- Фэрон-Демаре, Мюриэль (октябрь 1996 г.). « Dorinnotheca streelii Fairon-Demaret, gen. et sp. nov. , Новое раннее семенное растение из верхнего фамена Бельгии». Обзор палеоботаники и палинологии . 93 (1–4): 217–233. doi : 10.1016/0034-6667(95)00127-1 .
- Финни, ди-джей (ноябрь 1995 г.). «Фрэнк Йейтс, 12 мая 1902 г. - 17 июня 1994 г.». Биографические воспоминания членов Королевского общества . 41 : 554–573. doi : 10.1098/rsbm.1995.0033 . JSTOR 770162 . S2CID 26871863 .
- Флорос, Джон Д.; Ньюсом, Розетта; Фишер, Уильям (2010). «Накормить мир сегодня и завтра: важность пищевой науки и технологий» (PDF) . Институт пищевых технологов. Архивировано из оригинала (PDF) 16 февраля 2012 года . Проверено 1 марта 2012 г.
- Фрай, Южная Каролина (1989). «Структура и функции ксилоглюкана». Журнал экспериментальной биологии . 40 .
- Горд, Гордон; Хедрик, Д.Х. (2001). Словарь энтомологии . Кембридж, Массачусетс: Издательство CABI. ISBN 978-0-85199-291-4.
- Грей, Аса ; Сарджент, Чарльз (1889 г.). Научные статьи Асы Грея: выбрано Чарльзом Спрагом Сарджентом . Бостон, Массачусетс: Хоутон Миффлин . Проверено 26 февраля 2012 г.
- Грин, Эдвард Ли (1909). Вехи ботанической истории: изучение отдельных эпох в развитии науки ботаники: часть 1, До 1562 г. н.э. Вашингтон, округ Колумбия: Смитсоновский институт.
- Глинн, Джонатан М .; Миягисима, Шин-я; Йодер, Дэвид В .; Остерянг, Кэтрин В .; Вита, Станислав (1 мая 2007 г.). «Отдел хлоропластов» . Трафик . 8 (5): 451–461. doi : 10.1111/j.1600-0854.2007.00545.x . PMID 17451550 . S2CID 2808844 .
- Говик, У .; Вестхофф, П. (2010). «Путь от фотосинтеза C3 к фотосинтезу C4» . Физиология растений . 155 (1): 56–63. doi : 10.1104/стр.110.165308 . ПМС 3075750 . PMID 20940348 .
- Грайм, JP; Ходжсон, Дж. Г. (1987). «Ботанический вклад в современную экологическую теорию» . Новый фитолог . 106 (1): 283–295. doi : 10.1111/j.1469-8137.1987.tb04695.x . JSTOR 2433023 .
- Гаст, Девенс (1996). «Зачем изучать фотосинтез?» . Университет штата Аризона. Архивировано из оригинала 9 февраля 2012 года . Проверено 26 февраля 2012 г.
- Хэнкок, Джеймс Ф. (2004). Эволюция растений и происхождение видов сельскохозяйственных культур . Кембридж, Массачусетс: Издательство CABI. ISBN 978-0-85199-685-1.
- Хаберландт, Г. (1902). "Kulturverssuche mit isolierten Pflanzenzellen". Mathematicsch-naturwissenschaftliche (на немецком языке). 111 (1): 69–92.
- Харрис, Генри (2000). Рождение клетки . Нью-Хейвен, Коннектикут: Издательство Йельского университета. ISBN 978-0-300-08295-1.
- Хайнхорст, С .; Кэннон, ГК (январь 1993 г.). «Репликация ДНК в хлоропластах» . Журнал клеточной науки . 104 (104): 1–9. doi : 10.1242/jcs.104.1.1 . Проверено 2 июля 2013 г. .
- Эррера, см; Пеллмир, О. (2002). Взаимодействие растений и животных: эволюционный подход . Хобокен, Нью-Джерси: Blackwell Science. ISBN 978-0-632-05267-7.
- Хилл, Артур В. (1915). «История и функции ботанических садов» (PDF) . Анналы ботанического сада Миссури . 2 (1/2): 185–240. дои : 10.2307/2990033 . hdl : 2027/hvd.32044102800596 . JSTOR 2990033 .
- Хук, Кристиан; Манн, Д. Г.; Джанс, Х.М. (2005). Водоросли: Введение в Phycology . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-30419-1.
- Хоу, СиДжей; Барбрук, AC; Нисбет, RER; Локхарт, П.Дж.; Ларкум, AWD (2008). «Происхождение пластид» . Философские труды Королевского общества B: Биологические науки . 363 (1504): 2675–2685. doi : 10.1098/rstb.2008.0050 . ПВК 2606771 . PMID 18468982 .
- Хантер, Филип (май 2008 г.). «Что помнят гены» . Архивировано из оригинала 1 мая 2008 года . Проверено 24 августа 2013 г.
- Янзен, Дэниел Х. с рабочей группой завода CBOL; Форрест, Л. Л.; Спудж, Дж. Л.; Хаджибабаи, М.; и другие. (4 августа 2009 г.). «Штрих-код ДНК для наземных растений» . Труды Национальной академии наук . 106 (31): 12794–12797. Бибкод : 2009PNAS..10612794H . doi : 10.1073/pnas.0905845106 . ПВК 2722355 . PMID 19666622 .
- Ясечко, Скотт; Шарп, Закари Д.; Гибсон, Джон Дж.; Биркс, С. Джин; Йи, Йи; Фосетт, Питер Дж. (3 апреля 2013 г.). «Потоки наземных вод с преобладанием транспирации». Природа . 496 (7445): 347–350. Бибкод : 2013Natur.496..347J . doi : 10.1038/nature11983 . PMID 23552893 . S2CID 4371468 .
- Джеффрис, Диармуид (2005). Аспирин: Замечательная история чудо-лекарства . Нью-Йорк: Блумсбери. ISBN 978-1-58234-600-7.
- Джадд, штат Вашингтон; Кэмпбелл, CS; Келлог, Э.А.; Стивенс, ПФ; Донохью, MJ (2002). Систематика растений, филогенетический подход . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-403-4.
- Карп, Джеральд (2009). Клеточная и молекулярная биология: концепции и эксперименты . Хобокен, Нью-Джерси: John Wiley & Sons. ISBN 978-0-470-48337-4.
- Кенрик, Пол; Крейн, Питер Р. (сентябрь 1997 г.). «Происхождение и ранняя эволюция растений на суше». Природа . 389 (6646): 33–39. Бибкод : 1997Natur.389...33K . дои : 10.1038/37918 . S2CID 3866183 .
- Ким, Э .; Арчибальд, Дж. М. (2009). «Разнообразие и эволюция пластид и их геномов». В Санделиусе Анна Стина; Аронссон, Хенрик (ред.). Хлоропласт . Монографии клеток растений. Том. 13. CiteSeerX 10.1.1.325.3438 . doi : 10.1007/978-3-540-68696-5_1 . ISBN 978-3-540-68692-7.
- Клемм, Дитер; Хьюблен, Бриджит; Финк, Ханс-Петер; Бон, Андреас (6 сентября 2005 г.). «Целлюлоза: увлекательный биополимер и устойчивое сырье». ХимИнформ . 36 (36): 3358–93. doi : 10.1002/chin.200536238 . PMID 15861454 .
- Колаттукуди, Паппачан Э. (1996). "3" . В Kerstiens, G. (ред.). Кутикулы растений . Серия «Биология растений окружающей среды». Оксфорд: ISBN BIOS Scientific Publishers Ltd. 978-1-85996-130-8.
- Кресс, WJ; Вурдак, К.Дж.; Циммер, EA; Вейт, Луизиана; Янзен, Д.Х. (июнь 2005 г.). «Использование штрих-кодов ДНК для идентификации цветковых растений» . Труды Национальной академии наук . 102 (23): 8369–8374. Бибкод : 2005PNAS..102.8369K . doi : 10.1073/pnas.0503123102 . ПВК 1142120 . PMID 15928076 . Вспомогательная информация
- Ли, Эрнест К.; Сибриан-Харамильо, Анжелика; Колокотронис, Сергиос-Орестис; Катари, Манприт С .; Стаматакис, Александрос; Отт, Майкл; Чиу, Джоанна С .; Литтл, Дэймон П.; Стивенсон, Деннис В.; Маккомби, В. Ричард; Мартиенссен, Роберт А .; Коруцци, Глория; Десаль, Роб (2011). Сандерсон, Майкл Дж. (ред.). «Функциональный филогеномический взгляд на семенные растения» . ПЛОС Генетика . 7 (12): e1002411. doi : 10.1371/journal.pgen.1002411 . ПМС 3240601 . PMID 22194700 .
- Леонелли, Сабина; Чарнли, Беррис; Уэбб, Алекс; Бастоу, Рут (2012). «Под одним листом, исторический взгляд на Федерацию растениеводства Великобритании» . Новый фитолог . 195 (1): 10–13. doi : 10.1111/j.1469-8137.2012.4168.x . hdl : 10871/9251 . PMID 22530650 .
- Лепп, Хейно (2012). «Мхи» . Австралийский национальный ботанический сад . Проверено 14 июля 2013 г. .
- Леви, Мартин (1973). Ранняя арабская фармакология: введение на основе древних и средневековых источников . Лейден: Архив Брилла. ISBN 978-90-04-03796-0.
- Льюис, Луиза А.; Маккорт, Ричард М. (2004). «Зеленые водоросли и происхождение наземных растений». Американский журнал ботаники . 91 (10): 1535–1556. дои : 10.3732/ajb.91.10.1535 . PMID 21652308 . S2CID 14062252 .
- Лидделл, Генри Джордж; Скотт, Роберт (1940). Ботан (βοτάνη) . Оксфорд: Clarendon Press через цифровую библиотеку Perseus, Университет Тафтса.
- Лилберн, Тимоти Г .; Харрисон, Скотт Х .; Коул, Джеймс Р.; Гаррити, Джордж М. (2006). «Вычислительные аспекты систематической биологии» . Брифинги по биоинформатике . 7 (2): 186–195. doi : 10.1093/bib/bbl005 . PMID 16772262 .
- Лин, З .; Чжун, С .; Грирсон, Д. (2009). «Последние достижения в исследованиях этилена» . Журнал экспериментальной ботаники . 60 (12): 3311–3336. дои : 10.1093/jxb/erp204 . PMID 19567479 .
- Лопес-Баутиста, Х.М.; Уотерс, Д.А.; Чепмен, Р.Л. (2003). «Фрагмопластин, зеленые водоросли и эволюция цитокинеза» . Международный журнал систематической и эволюционной микробиологии . 53 (6): 1715–1718. doi : 10.1099/ijs.0.02561-0 . PMID 14657098 .
- Ланн, Дж. Э. (2002). «Эволюция синтеза сахарозы» . Физиология растений . 128 (4): 1490–500. doi : 10.1104/pp.010898 . ПМС 154276 . PMID 11950997 .
- Люттге, Ульрих (2006). «Фотосинтетическая гибкость и экофизиологическая пластичность: вопросы и уроки Clusia, единственного дерева CAM в Неотропах». Новый фитолог . 171 (1): 7–25. doi : 10.1111/j.1469-8137.2006.01755.x . JSTOR 3694480 . ПМИД 16771979 .
- Манн, Дж. (1987). Вторичный метаболизм, 2-е изд . Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-855529-2.
-
Мосет, Джеймс Д. (2003). Ботаника: Введение в биологию растений (3-е изд.). Садбери, Массачусетс: Обучение Джонса и Бартлетта. ISBN 978-0-7637-2134-3.
- Мосет, Джеймс Д. (2012). Ботаника: Введение в биологию растений (5-е изд.). Садбери, Массачусетс: Обучение Джонса и Бартлетта. ISBN 978-1-4496-6580-7.
- Макнил, Дж.; Барри, Франция; Бак, WR; Демулен, В.; Гройтер, В .; Хоксворт, DL; Херендин, PS; Кнапп, С.; Мархольд, К.; Прадо, Дж.; Прюдом Ван Рейн, WF; Смит, Г. Ф.; Вирсема, Дж. Х.; Терланд, Нью-Джерси (2011). Международный кодекс номенклатуры водорослей, грибов и растений (Мельбурнский кодекс), принятый Восемнадцатым международным ботаническим конгрессом, Мельбурн, Австралия, июль 2011 г. Том. Regnum Vegetabile 154. ARG Gantner Verlag KG. ISBN 978-3-87429-425-6.
- Медбери, шотландец (1993). «Таксономия и дизайн дендрария» (PDF) . Гарвардский университет . Проверено 26 июля 2013 г. .
- Митхила, Дж.; Холл, Дж. К.; Виктор, Дж. М.; Саксена, ПК (январь 2003 г.). «Тидиазурон индуцирует органогенез побегов при низких концентрациях и соматический эмбриогенез при высоких концентрациях на листовых и черешковых эксплантах африканской фиалки (Saintpaulia ionantha Wendl)». Отчеты о растительных клетках . 21 (5): 408–414. doi : 10.1007/s00299-002-0544-y . PMID 12789442 . S2CID 28072754 .
- Моргенсен, Х. Л. (1996). «Как и почему цитоплазматическое наследование у семенных растений». Американский журнал ботаники . 83 (3): 383–404. дои : 10.2307/2446172 . JSTOR 2446172 .
- Мортон, Алан Г. (1981). История ботанической науки: отчет о развитии ботаники с древних времен до наших дней . Лондон: Академическая пресса. ISBN 978-0-12-508380-5.
- Нидхэм, Джозеф; Лу, Гвей-джен; Хуанг, Син-Цунг (1986). Наука и цивилизация в Китае, Vol. 6 Часть 1 Ботаника . Кембридж: Издательство Кембриджского университета.
- Нобель, PS (1983). Биофизическая физиология растений и экология . Сан-Франциско: WH Freeman. ISBN 978-0-7167-1447-7.
- Оберлис, Томас (1998). Die Religion des Rgveda (на немецком языке). Вена: Sammlung De Nobili. ISBN 978-3-900271-31-2.
- Падманабхан, Мину С .; Динеш-Кумар, СП (2010). «Все руки на палубе - роль хлоропластов, эндоплазматического ретикулума и ядра в управлении врожденным иммунитетом растений» . Молекулярные взаимодействия растений и микробов . 23 (11): 1368–1380. doi : 10.1094/MPMI-05-10-0113 . PMID 20923348 .
- Панайно, Антонио (2002). Идеологии как межкультурные явления: материалы третьего ежегодного симпозиума проекта ассирийского и вавилонского интеллектуального наследия, состоявшегося в Чикаго, 27–31 октября 2000 г .. Болонья: Mimesis Edizioni. ISBN 978-88-8483-107-1.
- Порли, Рон; Ходжеттс, Ник (2005). Мхи и печеночники . Новая серия "Натуралист" №97 . Лондон: HarperCollins UK. ISBN 978-0-00-220212-1.
- Поссингхэм, СП; Роуз, Р. Дж. (18 мая 1976 г.). «Репликация хлоропластов и синтез ДНК хлоропластов в листьях шпината». Труды Королевского общества B: Биологические науки . 193 (1112): 295–305. Бибкод : 1976РССПБ.193..295П . doi : 10.1098/rspb.1976.0047 . S2CID 2691108 .
- Проктор, М.; Йео, П. (1973). Опыление цветов, серия «Новый натуралист» . Лондон: Харпер Коллинз. ISBN 978-0-00-219504-1.
- Рэйвен, Питер Х .; Эверт, Рэй Х .; Эйхорн, Сьюзен Э. (2005). Биология растений (7-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-1007-3.
- Рид, Ховард С. (1942). Краткая история наук о растениях . Нью-Йорк: Рональд Пресс.
- Рейк, Вольф (май 2007 г.). «Стабильность и гибкость регуляции эпигенетического гена в развитии млекопитающих». Природа . 447 (7143): 425–432. Бибкод : 2007Natur.447..425R . doi : 10.1038/nature05918 . PMID 17522676 . S2CID 11794102 .
- Реннер, СС; Риклефс, RE (1995). «Двудомность и ее корреляты у цветковых растений» (PDF) . Американский журнал ботаники . 82 (5): 596. doi : 10.2307/2445418 . JSTOR 2445418 .
- Роше, JD; Гольдшмидт-Клермон, М.; Торговец, Сабиха (1998). Молекулярная биология хлоропластов и митохондрий Chlamydomonas . Дордрехт, Германия: Kluwer Academic. ISBN 978-0-7923-5174-0.
- Ру, Стэнли Дж. (1984). «Действие Ca 2+ и фитохрома в растениях». Бионаука . 34 (1): 25–29. дои : 10.2307/1309422 . JSTOR 1309422 . PMID 11540810 .
- Руссин, Уильям А .; Эверт, Рэй Ф .; Вандервир, Питер Дж.; Шарки, Томас Д.; Бриггс, Стивен П. (1996). «Модификация определенного класса плазмодесм и потеря способности к экспорту сахарозы у мутанта кукурузы с дефектом экспорта 1» . Растительная клетка . 8 (4): 645–658. doi : 10.1105/tpc.8.4.645 . ПМС 161126 . PMID 12239395 .
- Сэттлер, Р. (1992). «Морфология процесса: структурная динамика в развитии и эволюции». Канадский журнал ботаники . 70 (4): 708–714. дои : 10.1139/b92-091 .
- Сэттлер, Р.; Жене, Б. (1992). «Многомерный анализ подтверждает континуум взгляда на форму растения». Анналы ботаники . 69 (3): 249–262. doi : 10.1093/oxfordjournals.aob.a088338 . JSTOR 42758718 .
- Савидан, Ю.Х. (2000). «Апомиксис: генетика и селекция». Обзоры селекции растений . 18 : 13–86. doi : 10.1002/9780470650158.ch2 . ISBN 978-0-470-65015-8.
- Шарф, Сара Т. (2009). «Ключи идентификации,« Естественный метод »и разработка руководств по идентификации растений». Журнал истории биологии . 42 (1): 73–117. doi : 10.1007/s10739-008-9161-0 . PMID 19831202 . S2CID 25763275 .
- Шарлеманн, JPW; Лоранс, ВФ (2008). «Насколько экологично биотопливо?». Наука . 319 (5859): 43–44. doi : 10.1126/science.1153103 . PMID 18174426 . S2CID 32144220 .
- Шелл, Дж.; Ван Монтегю, М. (1977). Ti-плазмида Agrobacterium tumefaciens – природный вектор для внедрения генов Nif в растения? . Основные науки о жизни . Том. 9. С. 159–179. doi : 10.1007/978-1-4684-0880-5_12 . ISBN 978-1-4684-0882-9. PMID 336023 .
- Шёнинг, Стив (2005). «Калифорнийский план действий по вредным и инвазивным сорнякам» (PDF) . Департамент продовольствия и сельского хозяйства Калифорнии. Архивировано из оригинала (PDF) 18 июля 2015 года . Проверено 1 марта 2012 г.
- Шнурр, Дж. А.; Шоки, Дж. М.; Де Бур, Г.Дж.; Обзор, JA (2002). «Экспорт жирных кислот из хлоропласта. Молекулярная характеристика основной пластидной ацил-коэнзима синтетазы из арабидопсиса» . Физиология растений . 129 (4): 1700–1709. doi : 10.1104/стр.003251 . ПВК 166758 . PMID 12177483 .
- Силин-Робертс, Хизер (2000). Письмо для науки и техники: документы, презентация . Оксфорд: Баттерворт-Хайнеманн. ISBN 978-0-7506-4636-9.
- Смолл, Майкл (2012). Динамика биологических систем . Бока-Ратон, Флорида: CRC Press. ISBN 978-1-4398-5336-8.
- Соботка, Роман; Сакова, Ленка; Курн, Владислав (2000). «Молекулярные механизмы самонесовместимости в капусте». Актуальные вопросы молекулярной биологии . 2 (4): 103–112. PMID 11471754 .
- Спектор, Тим (2012). Одинаково разные: почему вы можете изменить свои гены . Лондон: Вайденфельд и Николсон. ISBN 978-0-297-86631-2.
- Спраг, Т.А.; Спраг, MS (1939). «Травник Валериуса Кордуса». Журнал Линнеевского общества Лондона . ЛИИ (341): 1–113. doi : 10.1111/j.1095-8339.1939.tb01598.x .
- Стейс, Клайв А. (2010a). «Классификация по молекулам: что это даст полевым ботаникам?» (PDF) . Ватсония . 28 . Архивировано из оригинала (PDF) 26 июля 2011 г .. Проверено 06 июля 2013 г. .
- Стейс, Клайв (2010b). Новая флора Британских островов (3-е изд.). Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-70772-5.
- Старр, Сеси (2009). Единство и разнообразие жизни (изд. AP). Бельмомт, Калифорния: Брукс/Коул, Cenpage Learning. ISBN 978-1-111-58097-1.
- Стюарт, Уилсон Николс; Ротвелл, Гар В. (1993). Палеобиология и эволюция растений . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-38294-6.
- Стовер, Р. Х.; Симмондс, Северо-запад (1987). Бананы (3-е изд.). Харлоу, Англия: Лонгман. ISBN 978-0-582-46357-8.
- Самнер, Джудит (2000). Естественная история лекарственных растений . Нью-Йорк: Тимбер Пресс. ISBN 978-0-88192-483-1.
- Сун, Юх-Джу; Форухар, Фархад; Ли Хм, Хсоу-мин; Ту, Шух-Лонг; Йе, И-Хонг; Као, Сен; Шр, Хуэй-Линь; Чоу, Чиа-Чэн; Чен, Чинпан; Сяо, Чван-Дэн (2002). «Кристаллическая структура горохового Toc34, новой ГТФазы хлоропластного белка транслокона». Структурная биология природы . 9 (2): 95–100. дои : 10.1038/nsb744 . PMID 11753431 . S2CID 21855733 .
- Сассекс, И. (2008). «Научные корни современной биотехнологии растений» . Растительная клетка . 20 (5): 1189–1198. doi : 10.1105/tpc.108.058735 . ПВК 2438469 . PMID 18515500 .
- Таиз, Линкольн; Зейгер, Эдуардо (2002). Физиология растений (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-823-0.
- Такаичи, Шиничи (июнь 2011 г.). «Каротиноиды в водорослях: распределение, биосинтез и функции» . Морские наркотики . 9 (12): 1101–1118. дои : 10.3390/md9061101 . ПВК 3131562 . PMID 21747749 .
- Тэнсли, АГ (1935). «Использование и злоупотребление вегетативными терминами и понятиями». Экология . 16 (3): 284–307. дои : 10.2307/1930070 . JSTOR 1930070 .
- Тейлор, Теннесси; Тейлор, Э. Л.; Крингс, М. (2009). Палеоботаника, Биология и эволюция ископаемых растений (2-е изд.). Амстердам; Бостон: Академическая пресса. ISBN 978-0-12-373972-8.
- Томпсон, Джеймс Э .; Фрай, Стивен С. (2001). «Реструктуризация связанного со стенкой ксилоглюкана путем трансгликозилирования в клетках живых растений» . Журнал растений . 26 (1): 23–34. doi : 10.1046/j.1365-313x.2001.01005.x . PMID 11359607 . S2CID 18256045 .
- Ваггонер, Бен (2001). «Музей палеонтологии Калифорнийского университета» . Калифорнийский университет в Беркли . Проверено 27 февраля 2012 г.
- Пошел, FW; Тиманн, К.В. (1937). Фитогормоны (PDF) . Нью-Йорк: Макмиллан.
- Уиллис, Эй Джей (1997). «Экосистема: развивающаяся концепция с исторической точки зрения» . Функциональная экология . 11 (2): 268–271. doi : 10.1111/j.1365-2435.1997.00081.x .
- Везе, ЧР; Магрум, WE; Фокс, ЖЖ; Вулф, GE; Woese, RS (август 1977 г.). «Древнее расхождение среди бактерий». Журнал молекулярной эволюции . 9 (4): 305–311. Бибкод : 1977JMolE...9..305B . DOI : 10.1007/ BF01796092 . PMID 408502 . S2CID 27788891 .
- Янив, Зохара; Бахрах, Уриэль (2005). Справочник лекарственных растений . Бингемтон, Нью-Йорк: Haworth Press. ISBN 978-1-56022-994-0.
- Йейтс, Ф .; Мазер, К. (1963). «Рональд Эйлмер Фишер 1890–1962» . Биографические воспоминания членов Королевского общества . 9 : 91–129. doi : 10.1098/rsbm.1963.0006 . JSTOR 769423 .
- Зохари, Даниэль; Хопф, Мария (2000). Одомашнивание растений в Старом Свете (3-е изд.). Оксфорд: Издательство Оксфордского университета. ISBN 978-0-19-850356-9.
- Инициатива по геному арабидопсиса (2000 г.). «Анализ последовательности генома цветкового растения Arabidopsis thaliana» . Природа . 408 (6814): 796–815. Бибкод : 2000Natur.408..796T . дои : 10.1038/35048692 . PMID 11130711 .
- «Ауксины» . Гормоны растений, Исследовательская станция Лонг-Эштон, Исследовательский совет по биотехнологии и биологическим наукам . Проверено 15 июля 2013 г. .
- «Основное введение в науку, лежащую в основе ресурсов NCBI» . Национальный центр биотехнологической информации. 30 марта 2004 г. . Проверено 5 марта 2012 г.
- "Ботаника" . Интернет-словарь этимологии . 2012 . Проверено 24 февраля 2012 г.
- «Ранние травы - немецкие отцы ботаники» . Национальный музей Уэльса. 4 июля 2007 года. Архивировано из оригинала 29 июня 2012 года . Проверено 19 февраля 2012 г.
- «Екатерина Исава» . Национальный научный фонд. 1989 . Проверено 26 июня 2013 г.
- «Эволюция и разнообразие, ботаника для следующего тысячелетия: I. Интеллектуал: эволюция, развитие, экосистемы» . Ботаническое общество Америки . Проверено 25 июня 2013 г. .
- «Лекарство от трав» . Медицинский центр Мэрилендского университета. Архивировано из оригинала 02 марта 2012 г. Проверено 2 марта 2012 г.
- «Палеоботаника» . Кливлендский музей естественной истории . Проверено 30 июля 2014 г. .
- «Физическая карта брахиподиума » . Калифорнийский университет в Дэвисе. Архивировано из оригинала 24 августа 2011 года . Проверено 26 февраля 2012 г.
- «Растения и жизнь на Земле» . Ботанический сад Миссури. 2009 . Проверено 10 марта 2012 г.