
Гетероциста - Heterocyst

A – F: Nostoc commune G – H: Nostoc calcicola
I – M: Tolypothrix distorta N – R: Scytonema hyalinum
Шкала шкалы = 10 мкм, hc, гетероциста, ak, akinete, hm, гормогониум, nd, некридии
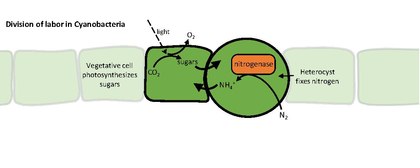
Гетероцисты или гетероциты - это специализированные азотфиксирующие клетки, образующиеся во время азотного голодания некоторыми нитчатыми цианобактериями , такими как Nostoc punctiforme , Cylindrospermum stagnale и Anabaena sphaerica . Они фиксируют азот из диазота (N 2 ) в воздухе с помощью фермента нитрогеназы , чтобы обеспечить клетки в нити азотом для биосинтеза.
Нитрогеназа инактивируется кислородом, поэтому гетероциста должна создавать микроанаэробную среду. Уникальная структура и физиология гетероцист требуют глобального изменения экспрессии генов . Например, гетероцисты:
- образуют три дополнительные клеточные стенки , в том числе одну из гликолипида, которая образует гидрофобный барьер для кислорода
- продуцируют нитрогеназу и другие белки, участвующие в азотфиксации
- разрушают фотосистему II , которая производит кислород
- до регулировать гликолитических ферментов
- производят белки, которые поглощают оставшийся кислород
- содержат полярные пробки, состоящие из цианофицина, который замедляет диффузию от клетки к клетке
Цианобактерии обычно получают фиксированный углерод (углевод) путем фотосинтеза . Отсутствие расщепления воды в фотосистеме II не позволяет гетероцистам выполнять фотосинтез, поэтому вегетативные клетки снабжают их углеводами , которые, как полагают, представляют собой сахарозу . Связанные источники углерода и азота обмениваются через каналы между ячейками в нити. Гетероцисты поддерживают фотосистему I , позволяя им генерировать АТФ путем циклического фотофосфорилирования .
Одиночные гетероцисты развиваются примерно через каждые 9-15 клеток, образуя одномерный узор вдоль филамента. Интервал между гетероцистами остается примерно постоянным, даже если клетки в филаменте делятся. Бактериальный филамент можно рассматривать как многоклеточный организм с двумя различными, но взаимозависимыми типами клеток. Такое поведение очень необычно для прокариот и, возможно, было первым примером формирования многоклеточного паттерна в эволюции . После того, как гетероциста сформировалась, она не может превратиться в вегетативную клетку. Некоторые бактерии, образующие гетероцисты, могут дифференцироваться в споровидные клетки, называемые акинетами, или подвижные клетки, называемые гормониями , что делает их наиболее фенотипически универсальными из всех прокариот.
Экспрессия гена

В среде с низким содержанием азота дифференцировка гетероцист запускается регулятором транскрипции NtcA. NtcA влияет на дифференцировку гетероцист с помощью сигнальных белков, участвующих в процессе дифференцировки гетероцист. Например, NtcA контролирует экспрессию нескольких генов, включая HetR, который имеет решающее значение для дифференцировки гетероцист. Это очень важно, поскольку он активирует другие гены, такие как hetR, patS, hepA, связываясь с их промотором и, таким образом, действуя как фактор транскрипции . Также стоит отметить, что экспрессия ntcA и HetR зависят друг от друга, и их присутствие способствует дифференцировке гетероцист даже в присутствии азота. Также недавно было обнаружено, что другие гены, такие как PatA, hetP, регулируют дифференцировку гетероцист. PatA формирует гетероцисты вдоль нитей, и он также важен для деления клеток . PatS влияет на формирование паттерна гетероцист путем ингибирования дифференцировки гетероцист, когда группа дифференцирующихся клеток объединяется, чтобы сформировать прогетероцисту (незрелую гетероцисту). Поддержание гетероцисты зависит от фермента hetN. Образование гетероцист ингибируется присутствием фиксированного источника азота , такого как аммоний или нитрат .
Образование гетероцисты
При образовании гетероцист из вегетативной клетки имеют место следующие последовательности:
- Клетка увеличивается в размерах.
- Гранулированные включения уменьшаются.
- Фотосинтетические переориентации ламелей.
- Стена окончательно становится трехслойной. Эти три слоя развиваются за пределами внешнего слоя клетки.
- Средний слой однородный.
- Внутренний слой ламинат.
- Стареющая гетероциста подвергается вакуолизации и, наконец, отрывается от нити, вызывая фрагментацию. Эти фрагменты называются гормогониями (единственное число Hormogonium ) и воспроизводятся бесполым путем.
Цианобактерии, образующие гетероцисты, делятся на отряды Nostocales и Stigonematales , которые образуют простые и ветвящиеся нити соответственно. Вместе они образуют монофилетическую группу с очень низкой генетической изменчивостью .
Симбиотические отношения
Бактерии также могут вступать в симбиотические отношения с некоторыми растениями. В таких отношениях бактерии реагируют не на доступность азота, а на сигналы, производимые растением для дифференцировки гетероцист. До 60% клеток могут стать гетероцистами, обеспечивая растение фиксированным азотом взамен фиксированного углерода. Сигнал, производимый растением, и стадия дифференцировки гетероцист, на которую оно влияет, неизвестны. Предположительно, симбиотический сигнал, генерируемый растением, действует до активации NtcA, поскольку hetR необходим для дифференцировки симбиотических гетероцист. Для симбиотической ассоциации с растением требуется ntcA, поскольку бактерии с мутировавшим ntcA не могут инфицировать растения.
Анабаена-Азолла
Заметные симбиотические отношения - это отношения цианобактерий Anabaena с растениями Azolla . Анабаена обитает на стеблях и в листьях растений азоллы . Azolla растений подвергаются фотосинтез и обеспечивает фиксированный углерод для Anabaena , чтобы использовать в качестве источника энергии для dinitrogenases в гетероцистах клеток. В свою очередь, гетероцисты способны обеспечивать вегетативные клетки и растение азолла фиксированным азотом в форме аммиака, который поддерживает рост обоих организмов.
Эти симбиотические отношения используются людьми в сельском хозяйстве. В Азии растения Azolla, содержащие виды Anabaena , используются в качестве биоудобрений в условиях ограниченного количества азота, а также в кормах для животных . Различные штаммы Azolla-Anabaena подходят для разных сред и могут приводить к различиям в урожайности сельскохозяйственных культур. Было показано, что рисовые культуры, выращенные с использованием азолла-анабаена в качестве биоудобрения, дают гораздо большее количество и качество продукции по сравнению с культурами без цианобактерий. Растения азолла-анабаена выращивают до и после посадки риса. По мере роста растения Azolla-Anabaena накапливают фиксированный азот из-за действия ферментов нитрогеназы и органического углерода в результате фотосинтеза растениями Azolla и вегетативными клетками Anabaena . Когда растения Azolla-Anabaena умирают и разлагаются, они выделяют большое количество фиксированного азота, фосфора , органического углерода и многих других питательных веществ в почву, обеспечивая богатую среду, идеально подходящую для роста рисовых культур.
Anabaena - Azolla отношение также было изучено в качестве возможного способа удаления загрязняющих веществ из окружающей среды, в процессе , известном как фиторемедиация . Anabaena sp. вместе с Azolla caroliniana успешно удаляют уран , токсичный загрязнитель, вызываемый горнодобывающей промышленностью , а также тяжелые металлы, ртуть (II) , хром (III) и хром (VI) из загрязненных сточных вод.

Растение Azolla caroliniana

Нить Anabaena circinalis

Цилиндроспермум филамент



использованная литература
- ^ Основы биологии (18 марта 2016 г.). «Бактерии» .
- ^ Wolk, CP; Эрнст, А .; Эльхай, Дж. (1994). Метаболизм и развитие гетероцист . Молекулярная биология цианобактерий . С. 769–823. DOI : 10.1007 / 978-94-011-0227-8_27 . ISBN 978-0-7923-3273-2.
- ^ Эрреро, Антония; Муро-пастор, Алисия М .; Флорес, Энрике (15 января 2001 г.). «Контроль азота в цианобактериях» . Журнал бактериологии . 183 (2): 411–425. DOI : 10.1128 / JB.183.2.411-425.2001 . ISSN 0021-9193 . PMC 94895 . PMID 11133933 .
- ^ Хига, Келли C .; Каллахан, Шон М. (1 августа 2010 г.). «Эктопическая экспрессия hetP может частично обходить потребность в hetR в дифференцировке гетероцист штаммом PCC 7120 Anabaena sp.» . Молекулярная микробиология . 77 (3): 562–574. DOI : 10.1111 / j.1365-2958.2010.07257.x . ISSN 1365-2958 . PMID 20545862 .
- ^ Ороско, Кристина С .; Рисер, Дуглас Д .; Каллахан, Шон М. (2006). «Анализ эпистаза четырех генов из штамма PCC 7120 Anabaena sp. Предполагает связь между PatA и PatS в формировании структуры гетероцист» . Журнал бактериологии . 188 (5): 1808–1816. DOI : 10.1128 / JB.188.5.1808-1816.2006 . ISSN 0021-9193 . PMC 1426565 . PMID 16484191 .
- ^ a b lee, Роберт Эдвард. Психология (PDF) . Проверено 9 октября 2017 года .
- ^ Микс, JC; Эльхай, Дж (2002). «Регулирование клеточной дифференцировки в нитчатых цианобактериях в свободноживущих и связанных с растениями симбиотических состояниях роста» . Обзоры микробиологии и молекулярной биологии . 66 (1): 94–121, содержание. DOI : 10.1128 / MMBR.66.1.94-121.2002 . PMC 120779 . PMID 11875129 .
- ^ a b c d van Hove, C .; Лежен, А. (2002). "Азолла: Симбиоз Анабаены". Биология и окружающая среда: материалы Королевской ирландской академии . 102B (1): 23–26. JSTOR 20500136 .
- ^ a b c d e f Vaishampayan, A .; Sinha, RP; Häder, D.-P .; Дей, Т .; Гупта, AK; Bhan, U .; Рао, А.Л. (2001). «Цианобактериальные биоудобрения в рисоводстве». Ботаническое обозрение . 67 (4): 453–516. DOI : 10.1007 / bf02857893 . JSTOR 4354403 .
- ^ Бокки, Стефано; Мальджоглио, Антонино (2010). «Azolla-Anabaenaas - биоудобрение для рисовых полей в долине реки По, рисовой зоне умеренного климата в Северной Италии» (PDF) . Международный журнал агрономии . 2010 : 1–5. DOI : 10.1155 / 2010/152158 . ISSN 1687-8159 .
- ^ Singh, S .; Prasad, R .; Singh, BV; Гоял, СК; Шарма, С. Н. (1990-06-01). «Влияние зеленых удобрений, сине-зеленых водорослей и мочевины, покрытой жмыхом нима, на рис с водно-болотных угодий (Oryza sativa L.)». Биология и плодородие почв . 9 (3): 235–238. DOI : 10.1007 / bf00336232 . ISSN 0178-2762 .
- ^ a b Bennicelli, R .; Stępniewska, Z .; Banach, A .; Szajnocha, K .; Островски, Дж. (2004-04-01). «Способность Azolla caroliniana удалять тяжелые металлы (Hg (II), Cr (III), Cr (VI)) из городских сточных вод». Chemosphere . 55 (1): 141–146. Bibcode : 2004Chmsp..55..141B . DOI : 10.1016 / j.chemosphere.2003.11.015 . PMID 14720557 .
- ^ Пан, Чанчунь; Ху, Нан; Дин, Дексин; Ху, Цзиньсонг; Ли, Гуанюэ; Ван, Ёндон (01.01.2016). «Экспериментальное исследование синергических эффектов между Азоллой и Анабаеной при удалении урана из растворов симбиотической системой Азолла-анабаена» . Журнал радиоаналитической и ядерной химии . 307 (1): 385–394. DOI : 10.1007 / s10967-015-4161-у . ISSN 0236-5731 .