
Тимидинкиназа - Thymidine kinase
| Тимидинкиназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллическая структура тетрамера тимидинкиназы из U. urealyticum (где мономеры имеют голубой, зеленый, красный и пурпурный цвета соответственно) в комплексе с тимидином ( модель заполнения пространства , углерод = белый, кислород = красный, азот = синий) .
| |||||||||
| Идентификаторы | |||||||||
| ЕС нет. | 2.7.1.21 | ||||||||
| № CAS | 9002-06-6 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
| Тимидинкиназа | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||
| Условное обозначение | ТЗ | ||||||||||
| Pfam | PF00265 | ||||||||||
| Клан пфам | CL0023 | ||||||||||
| ИнтерПро | IPR001267 | ||||||||||
| ПРОФИЛЬ | PDOC00524 | ||||||||||
| |||||||||||
| Тимидинкиназа 1, растворимая | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | TK1 | ||||||
| Ген NCBI | 7083 | ||||||
| HGNC | 11830 | ||||||
| OMIM | 188300 | ||||||
| RefSeq | NM_003258 | ||||||
| UniProt | P04183 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.7.1.21 | ||||||
| Locus | Chr. 17 q23.2-25.3 | ||||||
| |||||||
| Тимидинкиназа 2, митохондриальная | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Условное обозначение | TK2 | ||||||
| Ген NCBI | 7084 | ||||||
| HGNC | 11831 | ||||||
| OMIM | 188250 | ||||||
| RefSeq | NM_004614 | ||||||
| UniProt | O00142 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.7.1.21 | ||||||
| Locus | Chr. 16 [1] | ||||||
| |||||||
Тимидинкиназа - это фермент , фосфотрансфераза (киназа): 2'-дезокситимидинкиназа, АТФ-тимидин-5'-фосфотрансфераза, EC 2.7.1.21. Его можно найти в большинстве живых клеток. Он присутствует в клетках млекопитающих в двух формах, TK1 и TK2. Некоторые вирусы также обладают генетической информацией для экспрессии вирусных тимидинкиназ. Тимидинкиназа катализирует реакцию:
- Thd + ATP → TMP + ADP
где Thd представляет собой (дезокси) тимидин , ATP представляет собой аденозинтрифосфат , TMP представляет собой (дезокси) тимидинмонофосфат и ADP представляет собой аденозиндифосфат . Тимидинкиназы играют ключевую роль в синтезе ДНК и, следовательно, в делении клеток , поскольку они являются частью уникальной цепочки реакций по введению тимидина в ДНК. Тимидин присутствует в жидкостях организма в результате деградации ДНК из пищи и мертвых клеток. Тимидинкиназа необходима для действия многих противовирусных препаратов . Он используется для отбора линий гибридомных клеток для получения моноклональных антител . В клинической химии он используется в качестве маркера пролиферации при диагностике , контроле лечения и последующем наблюдении за злокачественными заболеваниями , в основном гематологическими злокачественными новообразованиями .
История
Включение тимидина в ДНК было продемонстрировано примерно в 1950 году. Несколько позже было показано, что этому включению предшествовало фосфорилирование , и примерно в 1960 году ответственный за это фермент был очищен и охарактеризован.
Классификация
Были идентифицированы два различных класса тимидинкиназ, которые включены в это суперсемейство: одно семейство объединяет тимидинкиназы из вируса герпеса, а также клеточные тимидилаткиназы, второе семейство группирует ТК из различных источников, включая позвоночных животных , бактерии , бактериофаг Т4. , поксвирусы , африканской чумы свиней вирус лихорадки (АЧС) и Фиша Lymphocystis вирусное заболевание (FLDV). Основной белок капсида радужных вирусов насекомых также принадлежит к этому семейству. Паттерн Prosite распознает только клеточный тип тимидинкиназ.
Изоферменты
У млекопитающих есть два изофермента , которые очень различаются по химическому составу, TK1 и TK2. Первый был впервые обнаружен в тканях плода, второй был обнаружен в большей степени в тканях взрослого человека, и первоначально они были названы тимидинкиназой плода и взрослого. Вскоре было показано, что TK1 присутствует в цитоплазме только в ожидании клеточного деления (зависит от клеточного цикла), тогда как TK2 находится в митохондриях и не зависит от клеточного цикла. Два изофермента имеют разную кинетику реакции и ингибируются разными ингибиторами.
Вирусные тимидинкиназы полностью отличаются от ферментов млекопитающих как структурно, так и биохимически и ингибируются ингибиторами, которые не ингибируют ферменты млекопитающих. Гены двух изоферментов человека были локализованы в середине 1970-х годов. Ген TK1 был клонирован и секвенирован. Соответствующий белок имеет молекулярную массу около 25 кДа. Обычно он встречается в ткани в виде димера с молекулярной массой около 50 кДа. Его можно активировать АТФ. После активации представляет собой тетрамер с молекулярной массой около 100 кДа. Однако форма фермента, присутствующего в кровотоке, не соответствует белку, кодируемому геном: основная фракция активного фермента в кровотоке имеет молекулярную массу 730 кДа и, вероятно, связана в комплексе с другими белками. Этот комплекс более стабилен и имеет более высокую удельную активность, чем любая из форм с более низкой молекулярной массой.
Рекомбинантный TK1 не может быть активирован и преобразован в тетрамер таким образом, что свидетельствует о том, что фермент, присутствующий в клетках, был модифицирован после синтеза.
TK1 синтезируется клеткой во время S-фазы клеточного деления. После завершения деления клеток TK1 разрушается внутриклеточно и не переходит в жидкости организма после нормального деления клеток. Существует обратная регуляция действия тимидинкиназы в клетке: тимидинтрифосфат (ТТФ), продукт дальнейшего фосфорилирования тимидина, действует как ингибитор тимидинкиназы. Это служит для поддержания сбалансированного количества ТТФ, доступного для синтеза нуклеиновых кислот, не перенасыщая систему. 5'-Аминотимидин, нетоксичный аналог тимидина, вмешивается в этот регуляторный механизм и тем самым увеличивает цитотоксичность аналогов тимидина, используемых в качестве противоопухолевых препаратов . В кинетика реакции тимидина и аналоги тимидина фосфорилирования является сложным и лишь частично известно. Общее фосфорилирование тимидина до тимидинтрифосфата не соответствует кинетике Михаэлиса-Ментен , а различные фосфаты тимидина и уридина препятствуют фосфорилированию друг друга. Кинетика TK от разных видов отличается друг от друга, а также разные формы от данного вида (мономер, димер, тетрамер и форма сыворотки) имеют разные кинетические характеристики.
Гены вирус-специфичных тимидинкиназ были идентифицированы в вирусе простого герпеса, вирусе ветряной оспы и вирусе Эпштейна-Барра.
 + АТФ --->
+ АТФ --->
 + АДФ
+ АДФ
Тимидин реагирует с АТФ с образованием монофосфата тимидина и АДФ.
Функция
Тимидинмонофосфат, продукт реакции, катализируемой тимидинкиназой, в свою очередь фосфорилируется до тимидиндифосфата с помощью фермента тимидилаткиназы и далее до тимидинтрифосфата с помощью фермента нуклеозиддифосфаткиназы . Трифосфат включен в молекулу ДНК, реакцию, катализируемую ДНК-полимеразой и комплементарной молекулой ДНК (или молекулой РНК в случае обратной транскриптазы , фермента, присутствующего в ретровирусе ).
Тимидина монофосфат также производится клеткой в другой реакции с помощью метилирования из дезоксиуридинмонофосфата , продукт других метаболических путей , не связанных с тимидина, под действием фермента тимидилатсинтазы . Второго пути достаточно для доставки тимидинмонофосфата для репарации ДНК. Когда клетка готовится к делению, требуется совершенно новая структура ДНК, и потребность в строительных блоках, включая тимидинтрифосфат, возрастает. Клетки готовятся к делению, вырабатывая некоторые ферменты, необходимые во время деления. Обычно они не присутствуют в клетках, и впоследствии их активность снижается, и они деградируют . Такие ферменты называются вспомогательными ферментами. Тимидинкиназа 1 является таким спасительным ферментом, тогда как тимидинкиназа 2 и тимидилатсинтаза не зависят от клеточного цикла.
Дефицит
Тимидинкиназа 2 используется клетками для синтеза митохондриальной ДНК. Мутации в гене TK2 приводят к миопатической форме синдрома истощения митохондриальной ДНК. Другой причиной дефицита TK2 может быть вызванное окислительным стрессом S-глутатионилирование и протеолитическая деградация митохондриальной тимидинкиназы 2. Синдром, вызванный дефицитом TK1, не известен, вероятно, поскольку дефектный ген TK1 может привести к гибели плода.
Тимидинкиназа во время развития
Образование тетрамера после модификации тимидинкиназы 1 после синтеза усиливает активность фермента. Было высказано предположение, что это механизм регуляции активности фермента. Образование тетрамеров наблюдается после стадии развития Dictyostelium. Предполагается, что его использование для тонкой регуляции синтеза ДНК было установлено у теплокровных животных после того, как они отошли от позвоночных. Также было изучено развитие тимидинкиназоподобных ферментов в развитии.
Распространение видов
Тимидинкиназа присутствует у животных, растений, некоторых бактерий, архей и вирусов. Были идентифицированы и в различной степени охарактеризованы тимидинкиназы из вирусов оспы, вируса африканской чумы свиней, вируса простого герпеса, вируса ветряной оспы и вируса Эпштейна-Барра. Форма фермента у вируса отличается от формы у других организмов. Тимидинкиназа отсутствует в грибах .
Приложения
Идентификация делящихся клеток
Первым непрямым использованием тимидинкиназы в биохимических исследованиях была идентификация делящихся клеток путем включения радиоактивно меченного тимидина и последующее измерение радиоактивности или авторадиография для идентификации делящихся клеток. Для этого в питательную среду добавляют меченный тритием тимидин. Несмотря на ошибки в методике, он все еще используется для определения скорости роста злокачественных клеток и изучения активации лимфоцитов в иммунологии.
ПЭТ-сканирование активных опухолей
Фтортимидин - аналог тимидина . Его захват регулируется тимидинкиназой 1, и поэтому он преимущественно поглощается быстро пролиферирующей опухолевой тканью. Изотоп фтора 18 представляет собой излучатель позитронов, который используется в позитронно-эмиссионной томографии (ПЭТ). Таким образом, радиоактивно меченый фтор-18 фтортимидин F-18 полезен для ПЭТ-визуализации активной пролиферации опухоли и выгодно отличается от более широко используемого маркера флюдезоксиглюкозы (18F) . Был предложен стандартизированный протокол, который поможет сравнить клинические исследования.
Подбор гибридом
Гибридомы - это клетки, полученные путем слияния опухолевых клеток (которые могут делиться бесконечно) и лимфоцитов, продуцирующих иммуноглобулин (плазматических клеток). Гибридомы могут быть расширены для производства большого количества иммуноглобулинов с заданной уникальной специфичностью (моноклональные антитела). Одна из проблем состоит в том, чтобы выделить гибридомы из большого избытка неслитых клеток после слияния клеток. Одним из распространенных способов решения этой проблемы является использование линий опухолевых клеток, отрицательных по тимидинкиназе (TK-), для слияния. Клетки, отрицательные по тимидинкиназе, получают путем выращивания линии опухолевых клеток в присутствии аналогов тимидина, которые убивают положительные по тимидинкиназе (TK +) клетки. Затем отрицательные клетки можно размножить и использовать для слияния с плазматическими клетками TK +. После слияния клетки выращивают в среде с метотрексатом или аминоптерином, которые ингибируют фермент дигидрофолатредуктазу, тем самым блокируя синтез тимидинмонофосфата de novo. Одной из таких обычно используемых сред является среда HAT, которая содержит гипоксантин, аминоптерин и тимидин. Неслитые клетки из линии клеток с дефицитом тимидинкиназы погибают, потому что они не имеют источника тимидинмонофосфата. Лимфоциты в конечном итоге умирают, потому что они не «бессмертны». Выживают только гибридомы, которые имеют «бессмертие» от своей клеточной линии-предка и тимидинкиназу от плазматической клетки. Затем отбирают те, которые продуцируют желаемое антитело, и культивируют для получения моноклонального антитела. Гибридомные клетки также могут быть выделены с использованием того же принципа, который описан в отношении другого гена, HGPRT, который синтезирует IMP, необходимый для синтеза нуклеотидов GMP в пути спасения.
Изучение структуры хромосом
Молекулярное расчесывание волокон ДНК можно использовать для отслеживания структуры хромосом у почкующихся дрожжей Saccharomyces cerevisiae. Это обеспечивает профили репликации ДНК отдельных молекул. Для этого необходимо, чтобы штаммы дрожжей экспрессировали тимидинкиназу, чего дрожжи дикого типа не экспрессируют, будучи грибами (см. «Вхождение»). Следовательно, ген тимидинкиназы должен быть включен в геном.
Клиническая химия
Тимидинкиназа - это спасательный фермент, который присутствует только в ожидании деления клеток. Фермент не высвобождается из клеток, подвергающихся нормальному делению, где у клеток есть особый механизм разрушения белков, которые больше не нужны после деления клетки. Поэтому у здоровых субъектов количество тимидинкиназы в сыворотке или плазме очень низкое. Опухолевые клетки выделяют фермент в кровоток, вероятно, в связи с разрушением мертвых или умирающих опухолевых клеток. Таким образом, уровень тимидинкиназы в сыворотке крови служит мерой злокачественной пролиферации, а косвенно - мерой агрессивности опухоли.
Терапевтические приложения
Некоторые препараты специально направлены против делящихся клеток. Их можно использовать против опухолей и вирусных заболеваний (как против ретровирусов, так и против других вирусов), поскольку больные клетки реплицируются намного чаще, чем нормальные клетки, а также против некоторых незлокачественных заболеваний, связанных с чрезмерно быстрой репликацией клеток (например, псориаза). Было высказано предположение, что противовирусная и противораковая активность аналогов тимидина, по крайней мере частично, достигается за счет подавления митохондриальной тимидинкиназы.
Цитостатики
Существуют различные классы лекарств, направленных против метаболизма тимидина и, таким образом, включающих тимидинкиназу, которые используются для контроля деления клеток, связанных с раком. Терминаторы цепи представляют собой аналоги тимидина, которые включены в растущую цепь ДНК, но модифицированы таким образом, чтобы цепь не удлинялась в дальнейшем. Как аналоги тимидина, препараты этого типа легко фосфорилируются до 5'-монофосфатов. Монофосфат далее фосфорилируется до соответствующего трифосфата и включается в растущую цепь ДНК. Аналог был модифицирован так, что он не имеет гидроксильной группы в 3'-положении, которое требуется для продолжения роста цепи. В зидовудине (AZT; ATC: J05AF01) 3'-гидроксильная группа заменена азидогруппой, в ставудине (ATC: J05AF04) она удалена без замены. AZT используется в качестве субстрата в одном из методов определения тимидинкиназы в сыворотке крови. Это означает, что АЗТ мешает этому методу и может быть ограничением: АЗТ является стандартным компонентом терапии ВААРТ при ВИЧ-инфекции. Одним из распространенных последствий СПИДа является лимфома, и наиболее важным диагностическим применением определения тимидинкиназы является мониторинг лимфомы.

AZT
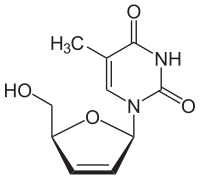
Ставудин

Идоксуридин

Ацикловир
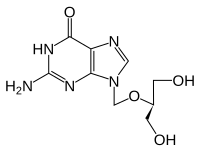
Ганцикловир





Другие аналоги тимидина , например идоксуридин (ATC: J05AB02), действуют, блокируя спаривание оснований во время последующих циклов репликации, тем самым делая полученную цепь ДНК дефектной. Это также можно комбинировать с радиоактивностью для достижения апоптоза злокачественных клеток.
Противовирусные препараты
Некоторые противовирусные препараты, такие как ацикловир (ATC: J05AB01) и ганцикловир (ATC: J05AB06), а также другие аналоги нуклеозидов используют субстратную специфичность вирусной тимидинкиназы в отличие от тимидинкиназ человека. Эти лекарства действуют как пролекарства, которые сами по себе не токсичны, но превращаются в токсичные лекарства путем фосфорилирования вирусной тимидинкиназой. Поэтому клетки, инфицированные вирусом, производят высокотоксичные трифосфаты, которые приводят к гибели клеток. Человеческая тимидинкиназа, напротив, с ее более узкой специфичностью, неспособна фосфорилировать и активировать пролекарство. Таким образом, к препарату чувствительны только клетки, инфицированные вирусом. Такие препараты эффективны только против вирусов из группы герпеса с их специфической тимидинкиназой. У пациентов, принимающих этот тип препаратов, часто наблюдается развитие устойчивости к противовирусным препаратам. Секвенирование гена тимидинкиназы в вирусе простого герпеса и вирусе ветряной оспы показывает быструю генетическую изменчивость и может облегчить диагностику устойчивости к противовирусным препаратам.
После того, как в декабре 1979 г. ВОЗ объявила о ликвидации оспы, программы вакцинации были прекращены. Повторное появление болезни случайно или в результате биологической войны затронет незащищенное население и может привести к эпидемии, которую будет трудно контролировать. Массовая вакцинация для борьбы с эпидемией оспы может быть сложной задачей, потому что единственная одобренная вакцина против оспы, вирус осповакцины, может иметь серьезные побочные эффекты. Тем не менее, некоторые правительства создают запасы вакцины против оспы, чтобы застраховаться от такой возможности. Однако приоритетной задачей является разработка конкретных и эффективных противовирусных препаратов. Одним из возможных подходов было бы использование для этой цели специфичности тимидинкиназы поксвируса, аналогично тому, как она используется для лекарств против вируса герпеса. Одна из трудностей состоит в том, что тимидинкиназа поксвируса принадлежит к тому же семейству тимидинкиназ, что и тимидинкиназы человека, и, следовательно, более похожа по химическому составу. Таким образом, структура тимидинкиназ поксвируса была определена для поиска потенциальных противовирусных препаратов. Однако поиски еще не привели к пригодному для использования противовирусному препарату против поксвирусов.
Как «суицидный ген» в генной терапии
Ген тимидинкиназы герпесвируса также использовался в качестве «суицидного гена» в качестве системы безопасности в экспериментах по генной терапии, позволяя убивать клетки, экспрессирующие ген, с помощью ганцикловира. Это желательно в случае, если рекомбинантный ген вызывает мутацию, приводящую к неконтролируемому росту клеток (инсерционный мутагенез). Цитотоксические продукты, продуцируемые этими модифицированными клетками, могут диффундировать к соседним клеткам, делая их так же восприимчивыми к ганцикловиру, явление, известное как «эффект свидетеля». Этот подход использовался для лечения рака на животных моделях, и его преимущество заключается в том, что опухоль может быть уничтожена всего лишь с помощью 10% злокачественных клеток, экспрессирующих ген. Аналогичная система была опробована с использованием тимидинкиназы томатов и AZT. Кроме того, ген тимидинкиназы используется в качестве суицидного гена для борьбы с опасной реакцией трансплантат против хозяина в терапии трансплантации гемопоэтических стволовых клеток под названием Zalmoxis, которая была условно одобрена в Европе в 2016 году.
Гены-маркеры опухолей
Подобное использование тимидинкиназы использует присутствие в некоторых опухолевых клетках веществ, не присутствующих в нормальных клетках ( опухолевые маркеры ). Такими опухолевыми маркерами являются, например, CEA (карциноэмбриональный антиген) и AFP (альфа-фетопротеин). Гены этих онкомаркеров можно использовать в качестве генов-промоторов тимидинкиназы. Затем тимидинкиназа может быть активирована в клетках, экспрессирующих опухолевый маркер, но не в нормальных клетках, так что лечение ганцикловиром убивает только опухолевые клетки. Однако такие основанные на генной терапии подходы все еще являются экспериментальными, поскольку проблемы, связанные с направленным переносом гена в опухолевые клетки, еще полностью не решены.
Нейтронозахватная терапия опухолей
Включение аналога тимидина с бором было предложено и опробовано на животных моделях для борной нейтронно-захватной терапии опухолей головного мозга. Описано очень большое количество производных тимидина, содержащих бор.
Противопаразитарные
Введение гена TK в геном паразита делает возможным включение BrdU и, таким образом, делает паразита чувствительным к лечению этим препаратом, что также было предложено и представляет собой чувствительный индикатор репликации генома паразита.
Измерение
В сыворотке и плазме
Уровни тимидинкиназы в сыворотке или плазме в основном измеряли с помощью анализов активности ферментов. В коммерческих анализах это делается путем инкубации образца сыворотки с аналогом субстрата и измерения количества образовавшегося продукта. Также использовалось прямое определение белка тимидинкиназы с помощью иммуноанализа. Количество тимидинкиназы, обнаруженное этим методом, плохо коррелирует с активностью фермента. Одна из причин этого заключается в том, что большое количество сывороточного TK1, идентифицированное иммуноанализом, не является ферментативно активным. Это особенно характерно для солидных опухолей, где иммуноанализы могут быть более чувствительными.
В ткани
Тимидинкиназа была определена в образцах ткани после извлечения ткани. Не было разработано ни одного стандартного метода экстракции или анализа, а определение ТК в экстрактах из клеток и тканей не было подтверждено в отношении какого-либо конкретного клинического вопроса, однако см. Romain et al. и Arnér et al. Разработан метод специфического определения TK2 в клеточных экстрактах с использованием аналога субстрата 5-бромовинил 2'-дезоксиуридина. В исследованиях, упомянутых ниже, использованные методы и способ представления результатов настолько различаются, что сравнения между разными исследованиями невозможны. Уровни TK1 в тканях плода во время развития выше, чем в соответствующих тканях позже. Некоторые незлокачественные заболевания также вызывают резкое повышение значений ТК в клетках и тканях: в периферических лимфоцитах при моноцитозе и в костном мозге при злокачественной анемии. Поскольку TK1 присутствует в клетках во время деления клеток, разумно предположить, что активность TK в злокачественной ткани должна быть выше, чем в соответствующей нормальной ткани. Это также подтверждается в большинстве исследований.
Иммуногистохимическое окрашивание
Антитела против тимидинкиназы доступны для иммуногистохимического определения. Окрашивание на тимидинкиназу оказалось надежным методом идентификации пациентов с карциномой молочной железы 2 стадии. Наибольшее количество идентифицированных пациентов было получено при сочетании окрашивания тимидинкиназы и Ki-67. Этот метод также был одобрен для лечения рака легких, колоректального рака, рака легких и почечно-клеточного рака.
Флуоресцентное окрашивание
2'-дезокси-2 ', 2'-дифтор-5-этинилуридин (dF-EdU) связывается с тимидинкиназой вируса простого герпеса, но из-за стерических затруднений не связывается с тимидинкиназой человека. Этот реагент вместе с флуоресцентным азидом вызывает флуоресценцию инфицированных клеток, но не неинфицированных. Следовательно, этот аналог субстрата позволяет специфически окрашивать инфицированные клетки.
Смотрите также
- Тимидинкиназа 1
- Тимидинкиназа вируса герпеса
- Тимидилаткиназа
- Нуклеозид-дифосфаткиназа
- Тимидилатсинтаза
- Тимидинкиназа в клинической химии
использованная литература
дальнейшее чтение
- Гароне С., Гарсиа-Диас Б., Эммануэле В., Лопес Л.С., Тадесс С., Акман Х.О. и др. (Август 2014 г.). «Обходная терапия дезоксипиримидинмонофосфатом при дефиците тимидинкиназы 2» . EMBO Молекулярная медицина . 6 (8): 1016–27. DOI : 10.15252 / emmm.201404092 . PMC 4154130 . PMID 24968719 .
- О'Нил К.Л., Баквалтер М.Р., Мюррей Б.К. (ноябрь 2001 г.). «Тимидинкиназа: диагностический и прогностический потенциал». Экспертный обзор молекулярной диагностики . 1 (4): 428–33. DOI : 10.1586 / 14737159.1.4.428 . PMID 11901857 . S2CID 12790327 .
- Тополькан О., Голубец Л. (февраль 2008 г.). «Роль тимидинкиназы при онкологических заболеваниях». Экспертное заключение по медицинской диагностике . 2 (2): 129–41. DOI : 10.1517 / 17530059.2.2.129 . PMID 23485133 .
- Джагарламуди К.К., Шоу М. (сентябрь 2018 г.). «Тимидинкиназа 1 как биомаркер опухоли: технический прогресс открывает новые возможности для старого биомаркера». Биомаркеры в медицине . 12 (9): 1035–1048. DOI : 10.2217 / ВММ-2018-0157 . PMID 30039979 .
внешние ссылки
- Тимидин + киназа в Национальных медицинских предметных рубриках США (MeSH)